

RSS-Feed abonnieren
DOI: 10.1055/a-1141-0151
Tara Tannin Regulates Pigmentation by Modulating Melanogenesis Enzymes and Melanosome Transport Proteins Expression
Autor*innen
Abstract
The skin color is imparted by the pigment melanin produced in the melanosomes of melanocytes, through the catalytic action of melanogenesis enzymes tyrosinase, tyrosinase-related protein 1, and dopachrome tautomerase. Disruptions in the melanogenesis process may result to hypopigmentation, as observed in cutaneous postinflammatory conditions. Here, the bioactivity of tara tannin, specifically on melanogenesis, was evaluated in vitro using human epidermal melanocytes (HEM) and B16F10 murine melanoma cells in order to determine the possibility that it may be used as a treatment against hypopigmentation. The melanin content of tara tannin-treated B16F10 cells and the expression level of melanogenesis enzymes and melanosome transport proteins were determined. To elucidate the underlying mechanism of tara tannin’s effect on melanogenesis, DNA microarray analysis was performed. Tara tannin significantly increased melanogenesis in both murine and human pigment cell models by upregulating melanogenesis-associated enzymes’ (tyrosinase, tyrosinase-related protein 1, and dopachrome tautomerase) protein and mRNA expression levels, as well as the melanosome transport proteins (myosin Va and RAB27A) expression, both attributed to increased microphthalmia-associated transcription factor (MITF) expression. Global gene expression analysis results revealed the modulation of genes (p≤0.05; fold-change ≥2.0 and ≤−2.0) that are under the transcriptional regulation of MITF and genes relevant for MAPK signaling, metabolic pathways, and cell cycle. Tara tannin has a significant effective melanogenesis-promoting effect, making it a potential therapeutic agent against hypopigmentation disorders. This is the first report on the melanogenesis regulatory effect of tara tannin in vitro.
Key words
melanogenesis - tara tannin - melanosome - Microphthalmia-Associated Transcription Factor (MITF)Introduction
Skin pigmentation contributes significantly to our overall appearance. A change in skin pigmentation, such what is observed in hypomelanosis, occurs when there is decreased melanogenesis or the failure of the mature melanosome to transfer to the dendritic tips of melanocytes [1]. Some of the diseases that cause hypomelanosis are chemical leukoderma, pityriasis alba, inflammatory diseases, and several infectious diseases [2]. Melanin is the biopolymer responsible for pigmentation and has important functions that include protection of the skin from UV radiation, neural cells from toxicants, and the inner ear from noise-induced temporary hearing loss [3] [4]. Moreover, melanin is strongly correlated with the prevention of the accumulation of free radicals/reactive oxygen species (ROS) generated by exposure of the skin to UV radiation [5]. Melanin is produced in the subcellular organelle melanosomes of neuro crest-derived melanocytes in a series of oxidation-reduction reactions catalyzed by melanogenic enzymes tyrosinase (TYR), tyrosinase-related protein 1 (TRP1), and dopachrome tautomerase (DCT) [6]. Effective pigmentation depends largely on the sorting and trafficking of melanosomes from melanocytes involving several proteins such as RAB27A and MYOVA, the so-called melanosome-transport proteins [7] [8] that, like the melanogenic enzymes, are under the transcriptional regulation of the microphthalmia-associated transcription factor (MITF) [9], the master regulator of melanogenesis [10]. In turn, several signaling pathways, including the cAMP-dependent pathway, regulate MITF [11].
In the past decade, there has been an increase in the reports on the efficacy of natural products in promoting melanogenesis [12] [13] [14]. These natural products, contained in plant extracts or as a pure compound, have been demonstrated to have a regulatory effect on MITF and melanogenesis by regulating either TYR expression or activity [15].
Tara tannin ([Fig. 1a]) is a natural product isolated from pods of tara (Caesalpinia spinosa) that has been extensively studied as a source of galloylquinic acids [16] and gallic acid [17]; except for its antifouling effect [18] [19], no other bioactivities of tara tannin have been reported. In this study, the effect of a tara tannin sample, which also includes other gallic acid derivatives and gallotannins ([Fig. 1b]), on melanogenesis was determined using the murine pigment cell model B16F10 cells and human epidermal melanocytes. To get an understanding of the signaling transduction or the mechanism underlying the observed effect on melanogenesis, the effect of tara tannin on the global gene expression in B16F10 cells was determined using DNA microarray.

Results
Before the effective concentration of tara tannin on melanogenesis was determined, the assessment of its cytotoxicity on B16F10 cells was determined first using the MTT assay. Results showed that treatment with 25, 50, and 75 µM tara tannin for 24 and 48 h did not have any cytotoxic effect on B16F10 cells ([Fig. 2a]). At 25 µM, cell proliferation actually increased significantly compared with the control group. Based on this result, tara tannin at 25 µM concentration was used in the succeeding experiments.

To determine the effect of tara tannin on melanin biosynthesis, the melanin content of B16F10 cells treated with 25 µM tara tannin or alpha melanocyte-stimulating hormone (α-MSH) for 48, 72, and 96 h was quantified. A significant increase in the intracellular melanin content (6-fold vs. control) was observed following treatment with tara tannin for 72 h ([Fig. 2b]). The increase in the melanin content was also evident in the pelleted melanin from cells treated with tara tannin for 72 h ([Fig. 2c]). It was also noted that after the 72 h treatment, the growth medium in 25 µM tara tannin-treated cells turned dark in color compared to the control and α-MSH plates ([Fig. 2d]) due to the melanosomes that were released from the cells. The total melanin produced by the cells, which included both the intracellular melanin and the melanin collected from the growth medium ([Fig. 2e]), was significantly increased by tara tannin.
To investigate the underlying reason for the increased melanogenesis, the effect of tara tannin on the expression of TYR, TRP1, and DCT was determined. Treatment with 25 µM tara tannin for 48 h increased the TYR, TRP1, and DCT expression, comparable to 200 nM α-MSH. ([Fig. 3a–c]). Results of the quantification of the mRNA expression level of these melanogenic enzymes showed that they were significantly upregulated, especially after 48 h of treatment ([Fig. 3]d–f). When the tara tannin-treated cells’ growth medium color was observed to have turned darker in color compared to the control, we hypothesized that it could be due to the melanin released from the cells, which means that tara tannin has an effect on melanosome transport. We then verified if our hypothesis was correct by determining the expression of melanosome transport-associated proteins MYO5A and RAB27A. As shown in [Fig. 3h] and [i], tara tannin increased MYO5A and RAB27A expression after 48 h. Since the melanogenic enzymes TYR, TRP1, and DCT, as well as the melanosome transport proteins MYO5A and RAB27A, are under the transcriptional regulation of MITF, determination of the effect of tara tannin on MITF expression was done, and the results showed that treatment with 25 µM tara tannin for 48 h increased the mRNA level of Mitf ([Fig. 3g]) and the MITF protein expression ([Fig. 3j]).

To find out if the observed effect of tara tannin on B16 cells will also be observed on human cells, the effect of tara tannin on human epidermal melanocytes (HEM) was established. First, noncytotoxic concentrations of tara tannin on HEM were determined using MTT assay, and the results showed that tara tannin was not cytotoxic at up to 10 µM. In contrast, there is a dose-dependent decrease in cell proliferation in mitomycin C-treated cells (positive control) as shown in [Fig. 4a]. HEM proliferation was actually increased when treated with 5 µM tara tannin ([Fig. 4a]). Based on this result, 5 µM tara tannin was used in the succeeding experiments. Quantification of the melanin content in HEM treated with 5 µM tara tannin or 200 nM α-MSH (positive control) showed that treatment with tara tannin for 72 h caused a 2-fold significant increase in the melanin content ([Fig. 4b]). The melanin content in [Fig. 4b] is the sum of the melanin present in the culture medium and the intracellular melanin ([Fig. 4c] and [d], respectively).
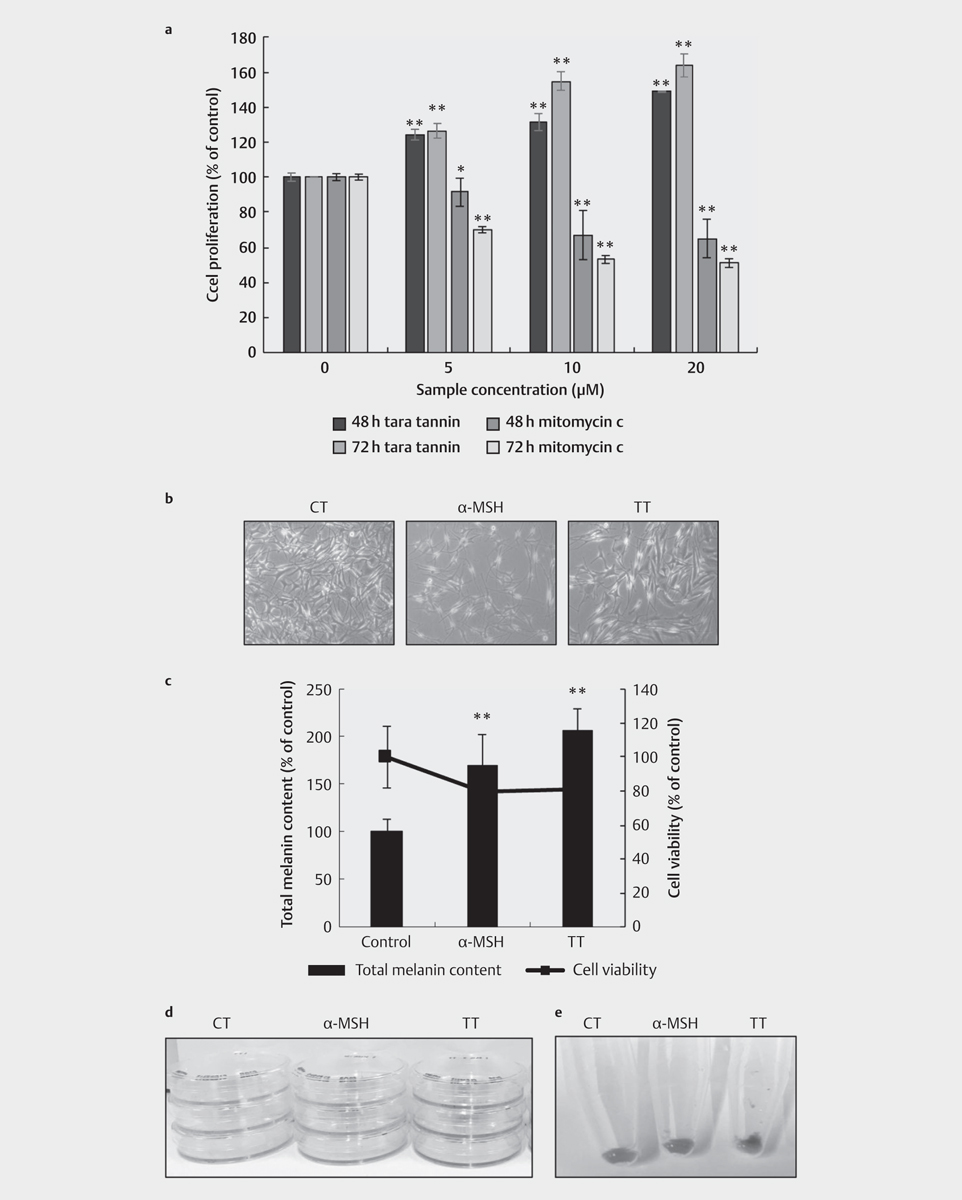
To investigate the underlying reason for the observed increase in melanogenesis in HEM following treatment with 5 µM tara tannin for 24, 48, and 72 h, the mRNA expression level of the melanogenic enzymes and MITF was quantified. Results showed that after 48 h treatment, 5 µM tara tannin significantly increased the TYR, TRP1, and DCT mRNA level ([Fig. 5a–c]) at all time points. Moreover, Mitf mRNA expression was also significantly upregulated by tara tannin treatment after 48 h (2.5-fold vs. control) and 72 h (1.5-fold vs. control). As expected, treatment with the hormone α-MSH increased the MITF expression by 1.5-fold (vs. control) ([Fig. 5d]). The blackish color of the cell culture medium was attributed to the release of melanin from the cells. The effect of tara tannin on melanosome transport was then evaluated by quantifying the mRNA level of MYO5A and RAB27A. Tara tannin upregulated RAB27A and MYO5A genes’ expression, with the increase in expression higher after 48 h of treatment than after 24 h and at its highest after 72 h ([Fig. 5e] and [f]).

To elucidate the molecular mechanisms affected by tara tannin treatment, the gene expression profile of B16F10 cells treated with tara tannin was determined. Differential expression analysis results showed that 1067 genes, out of which 720 genes were upregulated and 347 genes were downregulated, were modulated in B16F10 cells treated with tara tannin for 72 h. Significantly upregulated (p≥0.05) genes include Cxcr7, Tmem204, Fdft1, Cmpk2, Ankrd37, Mcm3, Adamts4, Narf, Aldoc, Dhcr24, Shisa2, Esco2, Insig1, Hmgcs1, Stard4, Ak4, Jarid2, Hspa1b, Tet1, and Nrep (Suppl. [Table 1a] ). The genes relevant for melanogenesis that were significantly expressed (fold-change ≥1.5 or ≤−1.5) were Kit, Prkca, Camk2g, Mitf, Prkaca, Plcb4, Mapk3, Adcy7, Fzd7, Ctnnb1, Map2k1, Adcy9, Adcy3, Fzd2, Fzd3, and Pomc (Suppl. [Table 1b]). Visualization of the modulated genes on Kegg Pathway, using DAVID, revealed that tara tannin-modulated genes that were significant in signaling pathways that regulate melanogenesis such as the MAPK and the cAMP signaling pathways ([Fig. 6]). The other genes that were differentially expressed by tara tannin treatment were significant in several pathways including the cell cycle, DNA replication, and metabolic pathways ([Table 1]). The number of genes that tara tannin modulated increased with the increase in treatment time (24 h, to 48 h, and to 72 h). Significant effect on melanogenesis-associated genes were observed after 72 h of treatment with tara tannin ([Fig. 6]).

|
Signaling pathways |
P-value |
|---|---|
|
Cell cycle |
2.7×10–13+ |
|
DNA replication |
7.1×10–11 |
|
Metabolic pathways |
2.5×10–10 |
|
Progesterone-mediated oocyte maturation |
2.4×10–9 |
|
Biosynthesis of antibiotics |
8.8×10–9 |
|
Oocyte meiosis |
3.3×10–7 |
|
Mismatch repair |
1.0×10–6 |
|
Steroid biosynthesis |
1.3×10–6 |
|
VEGF signaling pathway |
1.6×10–2 |
|
MAPK signaling pathway |
2.0×10–2 |
|
Melanogenesis |
3.0×10–2 |
1Analyzed using Transcriptome Analysis Console Software (Affymetrix).
Discussion
Skin and hair pigmentation are the most obvious human phenotypes, and a change in the color of some parts of the skin or hair causes severe psychological stress [1] [20]. We have previously reported that certain plant oil and extracts can promote melanogenesis [12] [14]. Here, we demonstrated how a plant-derived extract, tara tannin, promotes pigmentation. Tara tannin increased melanin biosynthesis as shown by the preliminary test results using murine pigment cell model B16F10 ([Figs. 2] and [3]). This melanogenesis promotion effect in murine cell model was also observed in HEM ([Fig. 4]), wherein the expression of melanogenic enzymes TYR, TRP1, and DCT as well as their transcription factor, MITF, were increased. Other natural products, such as quercetin, lupenone, and fisetin, have been reported to promote melanin biosynthesis by increasing the TYR activity [21] via the MAPK pathway [12] or by constitutive activation of CREB [22]. Similarly, tara tannin promotes melanogenesis but by increasing the melanogenesis enzyme TYR’s expression at the transcriptional level. At present, there are no reports on the effect of tara tannin on melanogenesis.
Tara tannin ([Fig. 1a]) is a main component of tara powder obtained from tara pods (Caesalpinia spinosa). It has a gallotannin structure [23] [24] and can thus be hydrolyzed enzymatically [25]. Tara powder has been reported to contain free gallic acid (2.6%) [26]. Even more gallic acid and ellagic acid may be released upon hydrolysis during, for example, prolonged storage). The tara powder used in this study also contained 2–3% gallic acid. Gallic acid and ellagic acid have inhibitory effect on melanogenesis [27] [28]. To evaluate the effect of gallic acid, B16 cells were treated with free gallic acid, and the results demonstrated that gallic acid also inhibits melanogenesis in our experimental setup (data not shown).
In this study, tara tannin-treated B16F10 or HEM cells promoted melanogenesis at up to 72 h of treatment ([Fig. 2b], [d], and [e]; [Fig. 4]b–[]d). It is also noteworthy to mention that tara tannin at lower concentration (5 μM) was effective in promoting melanogenesis in HEM while a higher dosage is required for murine cell model B16F10 wherein higher concentration appears to decrease the cell proliferation ([Fig. 4a]). Tara tannin modulates the signaling pathways ([Fig. 6]) that serve to regulate the expression of the genes of the enzymes catalyzing melanogenesis in B16F10 cells and in HEM ([Fig. 3]a–[]d; [Fig. 5]a–[]c) as well as upregulates MITF, the master regulator of melanogenesis ([Fig. 3e]; [Fig. 5d]). The expression of the melanogenic enzyme proteins B16F10 is not directly proportional to their observed mRNA expression, and this could be due to post-translational events upon which tara tannin may or may not have a direct effect.
As shown in [Fig. 2b], an increase in the melanin content was the highest (6-fold) in B16F10 cells treated with tara tannin for 72 h. This means that after 72 h, the melanin was “transported” or released from the cells (into the growth medium), and this could be the reason why the intracellular melanin content of the cells after the 96 h treatment is the same as the melanin content of the cells after 48 h ([Fig. 2e]). It is important to note that tara tannin can also promote melanosome transport since constitutive pigmentation is not just dependent on the quantity of melanin but also on the transfer and distribution from the melanocytes into the neighboring keratinocytes [7]. Moreover, the cellular organelle melanosome serves as the site of melanin synthesis, storage, and transport [8], and its uniform distribution in the epidermis characterizes normal pigmentation. Melanosomes are transported to the tips of the melanocyte dendrites via several melanosome transport proteins including MYOVA, RAB27A, and MLPH that bind to the melanosome [29]. Among these proteins, MYOVA and RAB27A play an important role in melanosome transport while melanophilin (MLPH) regulates the activity of MYOVA and dynein [30] [31]. Other reports also identify RAB27A and MLPH) as an organelle-associated receptor for MYOVA [29] [32]. In this study, tara tannin was shown to have a positive effect on melanosome transport, and this was supported by the data on the expression of the melanosome transport proteins MYOVA and RAB27 that were both increased by tara tannin ([Fig. 3]a, []b, [5]e and [f]) and demonstrated by the cell culture media becoming darker in color due to the presence of melanin ([Fig. 2d] and [Fig. 4c]). MITF is not just the master regulator of melanogenesis [33], but it is also the transcription factor of genes involved in melanocyte survival and several cellular events including melanosome transport [34] [35].
Using DNA microarray, tara tannin was found to modulate genes that are associated with melanogenesis (Suppl. [Table 1b]). Several signaling pathways that were also regulated by tara tannin as shown in [Table 1] (cell cycle, DNA replication, vascular endothelial growth factor) may not play a direct role in regulating melanogenesis, as illustrated in [Fig. 6], but may have contributed to the overall effect of tara tannin. The highly upregulated genes (Suppl. [Table 1a]) Cxcr7 regulates normal human epidermal melanocyte migration [36] while MCM3 is involved in the initiation of eukaryotic genome replication and shares a DNA replication factor with MITF [37]
An understanding of the genetic determinants of human pigmentation could help identify the molecular mechanisms of pigmentation-associated conditions including tanning response and skin cancers [38].
The benefits from using natural products in the stimulation of melanogenesis have long been recognized. Plants are a rich source of compounds that can promote melanogenesis [12] [14] [39] [40] and are one of the most widely used sources of pharmaceuticals. In this study, we have demonstrated that tara tannin can effectively increase the melanin production in B16F10 cells. This is the first report on the effect of tara tannin on melanogenesis and the underlying mechanism involved in this effect.
Materials and Methods
Cells and cell culture
B16F10 murine melanoma cells were purchased from the Riken Cell Bank in Tsukuba and cultured in RPMI1640 (Thermo Fisher Scientific) supplemented with 10% FBS (Sigma). Moderately pigmented neonatal HEM cells (Gibco) were cultured in Medium 254 (Thermo Fisher Scientific) supplemented with human melanocyte growth supplement with or without phorbol 12-meristate 13-acetate or PMA (HMGS) [S-016–5, (Thermo Fisher Scientific)]. Cells were incubated at 37°C in an incubator with 5% CO2. Photographs of the cell cultures were taken using Leica DMIL light microscope camera (DFC290 HD).
Tara tannin sample preparation and composition analysis
The tara tannin sample used in this study was extracted by Nano Innovation Laboratories, Ltd. from powdered Peruvian tara pods purchased from Kawamura Tsusho Co. Ltd. Briefly, a liter of ethanol was added to 0.33 kg tara powder, and the mixture was continuously stirred for 1 h at 60°C after which the mixture was cooled to 20°C and filtered in vacuo. Distilled water and ethanol were added and stirred (40°C). Finally, the solution was filtered in vacuo and stored in −20°C until use. The composition of this tara tannin sample was determined using HPLC with UV detector set at 254 nm. Samples were injected onto a TSKgel ODS-80Ts column (4.6φ × 150) (Tosoh Corporation) maintained at 40°C. A dual-gradient using acetonitrile/0.1% formic acid (solvent A) and water/0.1% formic acid (solvent B) was used for elution at a flow rate of 1 ml/min over 33 mins as follows: 95% B for 0.01 min; gradient to 65% B for 25 min; 5% B for 50 s min; gradient to 5% B for 1 min 10 s; 95% B for 2 min; and 95% for 3 min to re-equilibrate the column. For the bioassays, stock solution of tara tannin sample (>95% purity) was prepared by dissolving it in 70% ethanol (70% ethyl alcohol and 30% milli-Q water) and, prior to use, was filter-sterilized using a 0.22 μm filter (Merck Millipore) and stored at −20°C until use. Treatment with tara tannin was prepared by mixing the tara tannin stock solution in the growth medium for human epidermal melanocytes or B16F10 cells. Tara powder is rich in gallotannins and also contains small quantities of catechin derivatives. A characteristic compound is tara tannin (benzoic acid, 3,4-dihydroxy-5-((3,4,5-trihydroxybenzoyl)oxy)-, 5-(((4-carboxy-4-hydroxy-2,6-bis((3,4,5-trihydroxybenzoyl)oxy)cyclohexyl)oxy)carbonyl)-2,3-dihydroxyphenyl ester, (1S-(1-α,2-α,4-α,6-β))-) ([Fig. 1a]). HPLC-MS analysis revealed the presence of more than 40 individual gallotannins in tara pod extracts [41]. The tara powder sample used in this study showed a tannin fingerprint similar to that presented in ([Fig. 1b]) [41].
Cell viability assay
Briefly, B16F10 or HEM (3 × 103 cells/well) were seeded onto 96-well plates (Falcon) and incubated at 37oC in an incubator with 5% CO2. After overnight incubation, the growth medium was replaced by fresh growth medium with or without tara tannin (5, 10, 15, 25, 50, or 75 μM) then incubated for 24 and 48 h at 37oC in an incubator with 5% CO2. After specified incubation period, MTT solution at concentration 5 mg/ml (Wako) was added, and the cells incubated further for 6−8 h. To completely dissolve the formazan crystals, 10% sodium dodecyl sulfate (SDS) (Wako) was added, and incubated overnight. The cell viability was calculated based on the absorbances obtained at 570 nm using a microplate reader (Powerscan HT; Dainippon Pharmaceuticals USA Corporation). Blanks containing only medium, MTT, and SDS were subtracted from the average values of the absorbances. Mitomycin C (>95% purity; Merck) was used a positive control.
Melanin content determination
B16F10 cells or HEM were seeded at a density of 5 × 105 cells onto 10-cm petri dishes (Falcon) and cultured as described above. After 24 h incubation, the growth medium was replaced by fresh growth medium with or without 5, 10, 15, and 25 µM tara tannin or 200 nM α-MSH (Sigma;>95% purity), the positive control, and then incubated further for 48, 72, and 96 h. The cells were harvested by trypsination (0.25% trypsin⁄ 0.02% EDTA in PBS; Gibco) and solubilized by sonication after addition of 1% Triton X-100 (Sigma). Melanin was purified and precipitated in 10% trichloroacetic acid. Melanin in the spent growth medium was also collected for quantification. The precipitated melanin from both the cells and the growth medium was washed with 70% ethanol and then solubilized in 8 N NaOH with incubation at 80 °C for 2 h. The melanin content was then quantified spectrophotometrically (410 nm) and by comparing to a standard curve of synthetic melanin (Sigma) and expressed as melanin content per cell. The cell counts and cell viability were evaluated flow cytometrically as reported previously [14].
Western blot analysis
B16F10 cells (5 × 104 cells/ petri dishes) were seeded and incubated at 37°C in an incubator with 5% CO2. After 24 h incubation, the growth medium was replaced with fresh growth medium with or without 25 μM tara tannin and 200 nM α-MSH and incubated further for 24 and 48 h. After the specified incubation time, the protein samples were extracted using radio immunoprecipitation assay lysis buffer (Sigma) with 0.1% protease inhibitor cocktail (Sigma), loaded into 10% SDS-polyacrylamide gel, and subjected to electrophoresis (SDS-PAGE). The proteins were transferred onto PVDF membrane and incubated in specific primary antibodies against MITF (#110512; Assay Biotech); TYR (sc-7833; Santa Cruz Biotechnology; 1:200) goat polyclonal; TRP1 (sc-166857; Santa Cruz Biotechnology; 1:200) mouse monoclonal; DCT (sc-271356; Santa Cruz Biotechnology; 1:200) mouse monoclonal; RAB27A (sc-22756; Santa Cruz Biotechnology; 1:200) rabbit polyclonal; Myosin Va (sc-17706; Santa Cruz Biotechnology; 1:200) goat polyclonal; and GAPDH (sc-32233; Santa Cruz Biotechnology; 1:100) mouse monoclonal overnight at 4oC. Membranes were washed with PBS with Tween-20 (PBST) before incubation with goat antimouse IRDye 680LT or goat antiRabbit IRDye 800CW (LI-COR) secondary antibodies at room temperature. Detection was carried out using OdysseyFc Imaging System (LI-COR Inc.).
RNA extraction
B16F10 cells or HEM (5 × 104 cells/mL) were cultured as described above and the RNA extracted using ISOGEN (Nippon Gene) as previously reported [14]. The resulting RNA solution was quantified using Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific).
Real-time PCR
RNA samples (1μg) were reverse transcribed using the SuperScript III Reverse Transcription Kit (Invitrogen). The resulting cDNA was used as templates for real-time PCR (rt-PCR) using TaqMan Gene Expression Master Mix (Applied Biosystems), and specific primers for MITF (Hs01117294_m1), TYR (Hs00165976_m1), TRP1 (Hs00167051_m1), DCT (Hs0198278_m1), MYOVA (Hs00165309_m1), RAB27A (Hs00608302_m1), and GAPDH (Hs02786624_g1) (as an internal control) were used. Rt-PCR was performed using 7500 Fast Real-time PCR System with 7500 software version 2.0.5 (Applied Biosystems).
DNA microarray hybridization, imaging, and data analysis
DNA microarray was performed to determine the global gene expression changes in B16F10 cells in relation to the observed melanogenesis promotion effect of tara tannin. Single stranded cDNA was prepared from 200 ng of total RNA following the manufacturer’s instructions for Affymetrix Gene Chip 30 IVT Express Kit (Affymetrix). Total RNA was reverse transcribed into double-stranded cDNA, and biotin-labeled aRNA was generated using the 30 IVT Express Labeling Kit (Affymetrix). Biotin-labeled aRNA was hybridized to the Affymetrix mouse 430 PM Array strips (Affymetrix) for 16 h at 45°C at the Hybridization Station (Affymetrix). Hybridized arrays were washed and stained using the hybridization, wash, and stain Kit (Affymetrix) performed in Affymetrix GeneAtlasTM Fluidics Station. The arrays were scanned using the Affymetrix GeneAtlasTM Imaging Station.
The Affymetrix Expression Console Software was used to analyze the data by running comparisons of gene expression in treated and control cells based on mathematical algorithms. Results were based on the analysis of significance (control vs. treatment) using 1-way between-subject ANOVA (paired) (p value ≤0.05) and fold-change (linear) ≤ −2 or ≥2. The generated data was then analyzed using the Transcription Analysis Console Software. Gene ontology and functional annotation chart were derived using DAVID (database for annotation, visualization, and integrated discovery) (david.ncifcrf.gov). The DNA microarray data comply with MIAME guidelines and have been deposited in the ArrayExpress database at EMBL-EBI (www.ebi.ac.uk/arrayexpress).
Statistical analysis
Statistical analysis was carried out using the using Student’s t-test when 2 value sets were compared (control vs. sample). Mean values±standard error deviations (SD) were calculated, and a value of p≤0.05 was considered to be statistically significant.
For DNA microarray, the Affymetrix Expression Console Software was used to analyze the data by running comparisons of gene expression in treated and control cells based on mathematical algorithms. Results were based on the analysis of significance (control vs. treatment) using 1-way between-subject ANOVA (paired) (p value ≤0.05) and fold-change (linear) ≤−2 or ≥2.
Conflict of Interest
The authors declare that they have no conflict of interest.
Supporting information
- for this article is available online at
http://www.thieme-connect.de/products.
- Supporting information (PDF) (opens in new window)
-
References
- 1 Jy Lin, De Fisher. Melanocyte biology and skin pigmentation. Nature 2007; 445: 843-850
- 2 Hirobe T, Enami H. Melanoblast but not melanocytes decrease in number in human epidermis of idiopathic guttate hypomelanosis. Dermatol Sinica 2018; 36: 131-135
- 3 Ganesan AK, Ho H, Bodemann B, Petersen S, Aruri J, Koshy S, Richardson Z. et al. Genome-wide siRNA-based functional genomics of pigmentation identifies novel genes and pathways that impact melanogenesis in human cells. PLoS Genet 2008; 4: e1000298
- 4 Barrenäs M-L, Lindgren F. The influence of inner ear melanin on susceptibility to TTS in humans. Scand Audiol 1990; 19: 97-102
- 5 Herrling T, Jung K, Fuchs J. The role of melanin as protector against free radicals in skin and its role as free radical indicator in hair. Spectrochim Acta Part A Mol Biomol Spectrosc 2008; 69: 1429-1435
- 6 Hearing V, Tsukamoto K. Enzymatic control of pigmentation in mammals. FASEB J. 1991; 5: 2902-2909
- 7 Duval C, Chagnoleau C, Pouradier F, Sextius P, Condom E, Bernerd F. Human skin model containing melanocytes: essential role of keratinocyte growth factor for constitutive pigmentation—functional response to α-melanocyte stimulating hormone and forskolin. Tissue Eng Part C Methods 2012; 18: 947-957
- 8 Wasmeier C, Hume AN, Bolasco G, Seabra MC. Melanosomes at a glance. J Cell Sci 2008; 121: 3995-3999
- 9 Gaggioli C, Buscà R, Abbe P, Ortonne J-P, Ballotti R. Microphthalmia-associated transcription factor (MITF) is required but is not sufficient to induce the expression of melanogenic genes. Pigment Cell Res 2003; 16: 374-382
- 10 Steingrímsson E, Copeland NG, Jenkins NA. Melanocytes and the microphthalmia transcription factor network. Annu Rev Genet 2004; 38: 365-411
- 11 Khaled M, Larribere L, Bille K, Aberdam E, Ortonne JP, Balloti R, Bertolotto C. Glycogen synthase kinase 3beta is activated by cAMP and plays an active role in the regulation of melanogenesis. J Biol Chem 2002; 277: 33690-33697
- 12 Villareal MO, Han J, Matsuyama K, Sekii Y, Smaoui A, Shigemori H, Isoda H. Lupenone from Erica multiflora leaf extract stimulates melanogenesis in B16 murine melanoma cells through the inhibition of ERK1/2 activation. Planta Med. 2013; 79: 236-243
- 13 Matsuyama K, Villareal MO, El Omri A, Elyes Kchouk M, Han J, Isoda H. Effect of Tunisian Capparis spinosa L. extract on melanogenesis in B16 murine melanoma cells. J Nat Med 2009; 63: 468-472
- 14 Villareal MO, Kume S, Neffati M, Isoda H. Upregulation of Mitf by Phenolic compounds-rich cymbopogon schoenanthus treatment promotes melanogenesis in B16 melanoma cells and human epidermal melanocytes. BioMed Research International 2017; 2017: 1-11
- 15 Mo Villareal, Han J, Ikuta K, Isoda H. Mechanism of Mitf inhibition and morphological differentiation effects of hirsein A on B16 melanoma cells revealed by DNA microarray. J Dermatol Sci 2012; 67: 26-36
- 16 Clifford MN, Stoupi S, Kuhnert N. Profiling and characterization by LC-MSn of the galloylquinic acids of green tea, tara tannin, and tannic acid. J Agric Food Chem 2007; 55: 2797-2807
- 17 Deschamps AM, Lebeault J-M. Production of gallic acid from tara tannin by bacterial strains. Biotechnol Lett 1984; 6: 237-242
- 18 Bellotti N, del Amo B, Romagnoli R. Tara tannin a natural product with antifouling coating application. Progress Org. Coatings 2012; 74: 411-417
- 19 Bellotti N, del Amo B, Romagnoli R. Caesalpinia spinosa tannin derivaties for antifouling formulations. Procedia Materials Sci 2012; 1: 259-265
- 20 Hunt N, McHale S. The psychological impact of alopecia. BMJ Br Med J 2005; 331: 951-953
- 21 Nagata H, Takekoshi S, Takeyama R, Homma T, Yoshiyuki Osamura R. Quercetin enhances melanogenesis by increasing the activity and synthesis of tyrosinase in human melanoma cells and in normal human melanocytes. Pigment Cell Res 2004; 17: 66-73
- 22 Kumagai A, Horike N, Satoh Y, Uebi T, Sasaki T, Itoh Y, Hirata Y. et al. A potent inhibitor of SIK2, 3, 3′, 7-trihydroxy-4′-methoxyflavon (4′-o-methylfisetin), promotes melanogenesis in B16F10 melanoma cells. PLoS One 2011; 6: e26148
- 23 Haslam E, Hawroth D, Jones K, Rogers HJ. Gallotannins. Part 1. Introduction: and the fractionation of tannase. J Chem Soc 1961; 0: 1929-1935
- 24 Horler DF, Nursten HE. The tannins of tara, Caesalpinia spinosa (mol) kuntze. J Chem Soc 1961; 0: 3786-3792
- 25 Aouf C, Benyahya S, Esnouf A, Caillol S, Boutevin B, Fulcard H. Tara tannins as phenolic precursors of thermosetting apoxy. Eur Polymer J 2014; 55: 186-198
- 26 Garro Galvez JM, Riedl B, Conner AH. Analytical studies on tara tannins. Holzforschung 1997; 51: 235-243
- 27 Su TR, Lin JJ, Tsai CC, Huang TK, Yang ZY, Wu MO, Zheng YQ. et al. Inhibition of melanogenesis by gallic acid: possible involvement of the PI3K/Akt, MEK/ERK and Wnt/beta-catenin signaling pathways in B16F10. cells. J Mol Sci 2013; 14: 20443-20458
- 28 Shimogaki H, Tanaka Y, Tamai H, Masuda M. In vitro and in vivo evaluation of ellagic acid on melanogenesis inhibition. Int J Cosmet Sci 2000; 22: 291-303
- 29 Bahadoran P, Aberdam E, Mantoux F, Buscà R, Bille K, Yalman N, de Saint-Basile G. et al. Rab27a: A key to melanosome transport in human melanocytes. J Cell Biol 2001; 152: 843-850
- 30 Wu X, Bowers B, Rao K, Wei Q, Hammer JA. Visualization of melanosome dynamics within wild-type and dilute melanocytes suggests a paradigm for myosin V function in vivo . J Cell Biol 1998; 143: 1899-1918
- 31 Sheets L, Ransom DG, Mellgren EM, Johnson SL, Schnapp BJ. Zebrafish melanophilin facilitates melanosome dispersion by regulating dynein. Curr Biol 2007; 17: 1721-1734
- 32 Seabra MC, Coudrier E. RabGT Pases and myosin motors in organelle motility. Traffic 2004; 5: 393-399
- 33 García-Borrón JC, Sánchez-Laorden BL, Jiménez-Cervantes C. Melanocortin-1 receptor structure and functional regulation. Pigment Cell Res 2005; 18: 393-410
- 34 Chiaverini C, Beuret L, Flori E, Busca R, Abbe P, Bille K, Bahadoran P. et al. Microphthalmia-associated transcription factor regulates Rab27a gene expression and controls melanosome transport. J Biol Chem 2008; 283: 12635-12642
- 35 Vachtenheim J, Borovansky J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp Dermatol 2010; 19: 617-627
- 36 Lee E, Han J, Kim K, Choi H, Cho EG, Lee TR. CXCR7 mediates SDF1-induced melanocyte migration. Pigment Cell Melanoma Res 2013; 26: 58-66
- 37 Laurette P, Strub T, Koludrovic D, Keime C, Le Gras S, Seberg H, Van Otterloo E. et al. Transcription factor MITF and remodeller BRG1 define chromatin organisation at regulatory elements in melanoma cells. eLife 2015; 4: e06857
- 38 Han J, Kraft P, Nan H, Guo Q, Chen C, Qureshi A, Hankinson SE. et al. A genome-wide association study identifies novel alleles associated with hair color and skin pigmentation. PLoS Genet 2008; 4: e1000074
- 39 Villareal MO, Kume S, Neffati M, Isoda H. Upregulation of Mitf by phenolic compounds-rich cymbopogon schoenanthus treatment promotes melanogenesis in B16 melanoma cells and human epidermal melanocytes. BioMed Research International 2017; 2017: 1-11
- 40 Hong SH, Sim MJ, Kim YC. Melanogenesis-promoting effects of Rhynchosia nulubilis and Rhynchosia volubilis ethanol extracts in melan-a cells. Toxicol Res 2016; 32: 141-147
- 41 Venter P, Pasch H, de Villiers A. Comprehensive analysis of tara tannins by reversed-phase and hydrophilic interaction chromatography coupled to ion mobility and high-resolution mass spectrophotometry. Anal Bioanak Chem 2019; 411: 6329-6341
Correspondence
-
References
- 1 Jy Lin, De Fisher. Melanocyte biology and skin pigmentation. Nature 2007; 445: 843-850
- 2 Hirobe T, Enami H. Melanoblast but not melanocytes decrease in number in human epidermis of idiopathic guttate hypomelanosis. Dermatol Sinica 2018; 36: 131-135
- 3 Ganesan AK, Ho H, Bodemann B, Petersen S, Aruri J, Koshy S, Richardson Z. et al. Genome-wide siRNA-based functional genomics of pigmentation identifies novel genes and pathways that impact melanogenesis in human cells. PLoS Genet 2008; 4: e1000298
- 4 Barrenäs M-L, Lindgren F. The influence of inner ear melanin on susceptibility to TTS in humans. Scand Audiol 1990; 19: 97-102
- 5 Herrling T, Jung K, Fuchs J. The role of melanin as protector against free radicals in skin and its role as free radical indicator in hair. Spectrochim Acta Part A Mol Biomol Spectrosc 2008; 69: 1429-1435
- 6 Hearing V, Tsukamoto K. Enzymatic control of pigmentation in mammals. FASEB J. 1991; 5: 2902-2909
- 7 Duval C, Chagnoleau C, Pouradier F, Sextius P, Condom E, Bernerd F. Human skin model containing melanocytes: essential role of keratinocyte growth factor for constitutive pigmentation—functional response to α-melanocyte stimulating hormone and forskolin. Tissue Eng Part C Methods 2012; 18: 947-957
- 8 Wasmeier C, Hume AN, Bolasco G, Seabra MC. Melanosomes at a glance. J Cell Sci 2008; 121: 3995-3999
- 9 Gaggioli C, Buscà R, Abbe P, Ortonne J-P, Ballotti R. Microphthalmia-associated transcription factor (MITF) is required but is not sufficient to induce the expression of melanogenic genes. Pigment Cell Res 2003; 16: 374-382
- 10 Steingrímsson E, Copeland NG, Jenkins NA. Melanocytes and the microphthalmia transcription factor network. Annu Rev Genet 2004; 38: 365-411
- 11 Khaled M, Larribere L, Bille K, Aberdam E, Ortonne JP, Balloti R, Bertolotto C. Glycogen synthase kinase 3beta is activated by cAMP and plays an active role in the regulation of melanogenesis. J Biol Chem 2002; 277: 33690-33697
- 12 Villareal MO, Han J, Matsuyama K, Sekii Y, Smaoui A, Shigemori H, Isoda H. Lupenone from Erica multiflora leaf extract stimulates melanogenesis in B16 murine melanoma cells through the inhibition of ERK1/2 activation. Planta Med. 2013; 79: 236-243
- 13 Matsuyama K, Villareal MO, El Omri A, Elyes Kchouk M, Han J, Isoda H. Effect of Tunisian Capparis spinosa L. extract on melanogenesis in B16 murine melanoma cells. J Nat Med 2009; 63: 468-472
- 14 Villareal MO, Kume S, Neffati M, Isoda H. Upregulation of Mitf by Phenolic compounds-rich cymbopogon schoenanthus treatment promotes melanogenesis in B16 melanoma cells and human epidermal melanocytes. BioMed Research International 2017; 2017: 1-11
- 15 Mo Villareal, Han J, Ikuta K, Isoda H. Mechanism of Mitf inhibition and morphological differentiation effects of hirsein A on B16 melanoma cells revealed by DNA microarray. J Dermatol Sci 2012; 67: 26-36
- 16 Clifford MN, Stoupi S, Kuhnert N. Profiling and characterization by LC-MSn of the galloylquinic acids of green tea, tara tannin, and tannic acid. J Agric Food Chem 2007; 55: 2797-2807
- 17 Deschamps AM, Lebeault J-M. Production of gallic acid from tara tannin by bacterial strains. Biotechnol Lett 1984; 6: 237-242
- 18 Bellotti N, del Amo B, Romagnoli R. Tara tannin a natural product with antifouling coating application. Progress Org. Coatings 2012; 74: 411-417
- 19 Bellotti N, del Amo B, Romagnoli R. Caesalpinia spinosa tannin derivaties for antifouling formulations. Procedia Materials Sci 2012; 1: 259-265
- 20 Hunt N, McHale S. The psychological impact of alopecia. BMJ Br Med J 2005; 331: 951-953
- 21 Nagata H, Takekoshi S, Takeyama R, Homma T, Yoshiyuki Osamura R. Quercetin enhances melanogenesis by increasing the activity and synthesis of tyrosinase in human melanoma cells and in normal human melanocytes. Pigment Cell Res 2004; 17: 66-73
- 22 Kumagai A, Horike N, Satoh Y, Uebi T, Sasaki T, Itoh Y, Hirata Y. et al. A potent inhibitor of SIK2, 3, 3′, 7-trihydroxy-4′-methoxyflavon (4′-o-methylfisetin), promotes melanogenesis in B16F10 melanoma cells. PLoS One 2011; 6: e26148
- 23 Haslam E, Hawroth D, Jones K, Rogers HJ. Gallotannins. Part 1. Introduction: and the fractionation of tannase. J Chem Soc 1961; 0: 1929-1935
- 24 Horler DF, Nursten HE. The tannins of tara, Caesalpinia spinosa (mol) kuntze. J Chem Soc 1961; 0: 3786-3792
- 25 Aouf C, Benyahya S, Esnouf A, Caillol S, Boutevin B, Fulcard H. Tara tannins as phenolic precursors of thermosetting apoxy. Eur Polymer J 2014; 55: 186-198
- 26 Garro Galvez JM, Riedl B, Conner AH. Analytical studies on tara tannins. Holzforschung 1997; 51: 235-243
- 27 Su TR, Lin JJ, Tsai CC, Huang TK, Yang ZY, Wu MO, Zheng YQ. et al. Inhibition of melanogenesis by gallic acid: possible involvement of the PI3K/Akt, MEK/ERK and Wnt/beta-catenin signaling pathways in B16F10. cells. J Mol Sci 2013; 14: 20443-20458
- 28 Shimogaki H, Tanaka Y, Tamai H, Masuda M. In vitro and in vivo evaluation of ellagic acid on melanogenesis inhibition. Int J Cosmet Sci 2000; 22: 291-303
- 29 Bahadoran P, Aberdam E, Mantoux F, Buscà R, Bille K, Yalman N, de Saint-Basile G. et al. Rab27a: A key to melanosome transport in human melanocytes. J Cell Biol 2001; 152: 843-850
- 30 Wu X, Bowers B, Rao K, Wei Q, Hammer JA. Visualization of melanosome dynamics within wild-type and dilute melanocytes suggests a paradigm for myosin V function in vivo . J Cell Biol 1998; 143: 1899-1918
- 31 Sheets L, Ransom DG, Mellgren EM, Johnson SL, Schnapp BJ. Zebrafish melanophilin facilitates melanosome dispersion by regulating dynein. Curr Biol 2007; 17: 1721-1734
- 32 Seabra MC, Coudrier E. RabGT Pases and myosin motors in organelle motility. Traffic 2004; 5: 393-399
- 33 García-Borrón JC, Sánchez-Laorden BL, Jiménez-Cervantes C. Melanocortin-1 receptor structure and functional regulation. Pigment Cell Res 2005; 18: 393-410
- 34 Chiaverini C, Beuret L, Flori E, Busca R, Abbe P, Bille K, Bahadoran P. et al. Microphthalmia-associated transcription factor regulates Rab27a gene expression and controls melanosome transport. J Biol Chem 2008; 283: 12635-12642
- 35 Vachtenheim J, Borovansky J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp Dermatol 2010; 19: 617-627
- 36 Lee E, Han J, Kim K, Choi H, Cho EG, Lee TR. CXCR7 mediates SDF1-induced melanocyte migration. Pigment Cell Melanoma Res 2013; 26: 58-66
- 37 Laurette P, Strub T, Koludrovic D, Keime C, Le Gras S, Seberg H, Van Otterloo E. et al. Transcription factor MITF and remodeller BRG1 define chromatin organisation at regulatory elements in melanoma cells. eLife 2015; 4: e06857
- 38 Han J, Kraft P, Nan H, Guo Q, Chen C, Qureshi A, Hankinson SE. et al. A genome-wide association study identifies novel alleles associated with hair color and skin pigmentation. PLoS Genet 2008; 4: e1000074
- 39 Villareal MO, Kume S, Neffati M, Isoda H. Upregulation of Mitf by phenolic compounds-rich cymbopogon schoenanthus treatment promotes melanogenesis in B16 melanoma cells and human epidermal melanocytes. BioMed Research International 2017; 2017: 1-11
- 40 Hong SH, Sim MJ, Kim YC. Melanogenesis-promoting effects of Rhynchosia nulubilis and Rhynchosia volubilis ethanol extracts in melan-a cells. Toxicol Res 2016; 32: 141-147
- 41 Venter P, Pasch H, de Villiers A. Comprehensive analysis of tara tannins by reversed-phase and hydrophilic interaction chromatography coupled to ion mobility and high-resolution mass spectrophotometry. Anal Bioanak Chem 2019; 411: 6329-6341