

Subscribe to RSS
DOI: 10.1055/s-0035-1557833
Assessment of the Effect of Arbutin Isomers and Kojic Acid on Melanin Production, Tyrosinase Activity, and Tyrosinase Expression in B16-4A5 and HMV-II Melanoma Cells
Authors
Correspondence
Publication History
received 14 March 2015
revised 04 June 2015
accepted 18 July 2015
Publication Date:
09 September 2015 (online)
Abstract
The inhibitory effects of α-arbutin, β-arbutin, and kojic acid on melanogenesis, tyrosinase activity, and tyrosinase protein expression in mouse melanoma cells (B16-4A5) and human melanoma cells (HMV-II) were directly compared. β-Arbutin showed a stronger inhibitory effect on melanogenesis and tyrosinase expression in B16-4A5 cells than α-arbutin and kojic acid. Kojic acid showed a stronger inhibitory effect on mushroom and B16-4A5 tyrosinase activity than α-arbutin and β-arbutin. In contrast, kojic acid inhibited all of these effects more strongly than α-arbutin or β-arbutin in HMV-II cells. These results suggest that kojic acid may be used as a positive control for the inhibitory melanogenesis assay, and for tyrosinase activity and tyrosinase expression assays that use HMV-II cells. Moreover, using HMV-II cells with kojic acid as the positive control may facilitate the search for new skin-whitening agents using natural products and provide an alternative to the B16-4A5 assay.
The production of excessive pigmentation in melasma, spots, freckles, and other forms of hyperpigmentation can pose a significant aesthetic problem [1], [2]. The causative pigment, melanin, is formed by a combination of enzymatically catalyzed chemical reactions. Melanogenesis is initiated by the catalytic oxidation of tyrosine to dopaquinone by tyrosinase; this is the rate-limiting step in melanin synthesis [3]. Tyrosinase inhibitors reduce or block melanin synthesis, leading to skin whitening. Several common skin-lightening and depigmentation agents available commercially include arbutin (hydroquinone β-D-glucopyranoside, β-ARB), kojic acid (KA), and hydroquinone [4], [5]. These compounds are isolated from natural resources and are bioassayed using mushroom tyrosinase and B16 mouse melanoma cells. In a recent study, human melanoma HMV-II cells were used as an alternative to B16 [6], [7]. α-Arbutin (α-ARB), the epimer of β-ARB ([Fig. 1]), has inhibitory effects on melanogenesis and tyrosinase [8], [9]. However, it remains unclear whether β-ARB or α-ARB has strong activity against HMV-II, and the inhibitory effects of KA on melanogenesis and tyrosinase using HMV-II are also unclear. In this study, we directly compared the inhibitory effects of these compounds on melanogenesis and tyrosinase activity using B16 and HMV-II.

The inhibitory effect of α-ARB, β-ARB, and KA on the proliferation and pigmentation of B16-4A5 and HMV-II were compared. [Fig. 2 A] shows that α-ARB, β-ARB, and KA at 125 to 1000 µM suppressed melanogenesis in B16-4A5. β-ARB significantly suppressed melanogenesis in a concentration-dependent manner between 125 and 1000 µM, α-ARB significantly suppressed melanogenesis in a concentration-dependent manner between 250 and 1000 µM, whereas KA only significantly suppressed melanogenesis at 1000 µM. These results showed that β-ARB suppressed melanogenesis more effectively than α-ARB and KA in B16-4A5. [Fig. 2 B] shows that α-ARB, β-ARB, and KA at 125 to 1000 µM suppressed melanogenesis in HMV-II, KA significantly suppressed melanogenesis in a concentration-dependent manner between 125 and 1000 µM, whereas α-ARB and β-ARB significantly suppressed melanogenesis in a concentration-dependent manner between 500 and 1000 µM. These results demonstrated that KA was superior compared to α-ARB and β-ARB at suppressing melanogenesis in HMV-II. The inhibitory effects of these compounds on melanogenesis in HMV-II were, thus, in contrast with their inhibitory effects on melanogenesis in B16-4A5. Significant cytotoxic effects were only observed for α-ARB at 1000 µM on B16-4A5, β-ARB at 500 and 1000 µM on B16-4A5, and β-ARB at 1000 µM on HMV-II (Fig. 1 S, Supporting Information).

The inhibition of tyrosinase or/and downregulation of tyrosinase expression are mechanisms by which depigmentation occurs [3], [10]. Mushroom tyrosinase, B16-4A5, and HMV-II are used to screen for inhibitors of tyrosinase and suppressors of melanin biosynthesis [3], [4], [11]. [Table 1] shows the inhibitory effect of α-ARB, β-ARB, and KA on mushroom, B16-4A5, and HMV-II tyrosinase. α-ARB interestingly showed an inhibitory effect on B16-4A5 tyrosinase (297.4 ± 9.7 µM) but no inhibitory effect on mushroom and HMV-II tyrosinases (> 500 µM). In contrast, β-ARB showed weak inhibition of all three tyrosinases (> 500 µM). The KA IC50 inhibition values of mushroom, B16-4A5, and HMV-II tyrosinases were 297.4 ± 9.7 µM, 57.8 ± 1.8 µM, and 223.8 ± 4.9 µM, respectively.
|
Mushroom tyrosinase |
B16-4A5 tyrosinase |
HMV-II tyrosinase |
|
|---|---|---|---|
|
Values for each compound represent the mean ± SD of four independent tests |
|||
|
α-ARB |
> 500 |
297.4 ± 9.7 |
> 500 |
|
β-ARB |
> 500 |
> 500 |
> 500 |
|
KA |
182.7 ± 9.8 |
57.8 ± 1.8 |
223.8 ± 4.9 |
We postulated that the melanogenesis inhibitory effect of KA might have contributed to the inhibition of tyrosinase. To investigate the mechanism by which α-ARB, β-ARB, and KA inhibit melanogenesis, we used Western blotting to examine the effect of these compounds on the expression of B16-4A5 and HMV-II tyrosinase proteins ([Fig. 3]). In B16-4A5, tyrosinase expression was decreased by β-ARB, α-ARB, and KA (all 500 µM, listed in order of decreasing effect) and this ordering of the compounds is the same as the order of their effect on reducing melanin in B16-4A5. In HMV-II, tyrosinase expression was decreased by KA, β-ARB, and α-ARB (all 1000 µM, listed in order of decreasing effect), which is the same order as their effect on reducing melanin content. KA is a known tyrosinase inhibitor, but it does not decrease tyrosinase expression in several B16 cell lines [12]. However, there are no previous reports directly comparing the effect of KA with other agents in decreasing tyrosine levels in HMV-II, as apposed to B16 tyrosinase levels. The present data confirmed that the inhibition of melanogenesis by KA is due to a specific inhibition and downregulation of HMV-II tyrosinase.
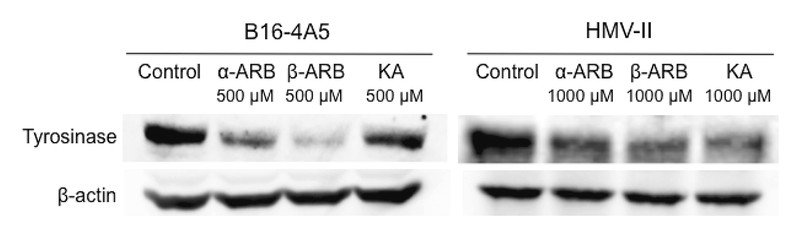
In conclusion, KA directly inhibited tyrosinase activity and suppressed the expression levels of tyrosinase in HMV-II. Furthermore, this is the first report to directly compare the effects of α-ARB, β-ARB, and KA on melanogenesis and to evaluate their inhibitory effect on tyrosinase in B16-4A5 and HMV-II. These results suggest that KA can be used as a positive control in the assessment of melanogenesis, tyrosinase activity, and tyrosinase expression in HMV-II cells, thereby providing an alternative to the mushroom tyrosinase assay and B16-4A5 assay in the search for new skin-whitening agents derived from natural products ([Table 2]).
|
Mushroom |
B16-4A5 |
HMV-II |
|
|---|---|---|---|
|
Melanin production |
– |
β-ARB |
KA |
|
Tyrosinase activity |
KA |
KA |
KA |
|
Tyrosinase expression |
– |
β-ARB |
KA |
Materials and Methods
Cell cultures: B16-4A5 (RCB0557) and HMV-II (RCB0777) cells were obtained from the RIKEN BRC through the National Bio-Resource Project of the MEXT, Tsukuba, Japan. B16-4A5 cells were cultured in DMEM supplemented with 10 % fetal bovine serum (FBS; Nichirei Biosciences), penicillin, and streptomycin at 37 °C in a humidified atmosphere consisting of 5 % CO2 and 95 % air. HMV-II cells were cultured in Hamʼs F12 supplemented with 10 % FBS (Vitromex), penicillin, and streptomycin at 37 °C in a humidified atmosphere consisting of 5 % CO2 and 95 % air.
Measurement of melanin content: The amount of melanin in cultured melanoma cells was measured as described previously [13], [14]. See Supporting Information for detailed protocols.
Tyrosinase inhibition assay: Mushroom tyrosinase activity was measured by determining its DOPA-oxidase activity using a modification of the method of Ha et al. [15]. First, 120 µL of L-DOPA (8 mM, dissolved in 50 mM phosphate buffer, pH 6.8) and 40 µL of the sample were mixed. Then, 40 µL of mushroom tyrosinase (80 units/mL) was added, and the amount of dopachrome in the reaction mixture was determined by measuring the optical density at 492 nm after 20 min at 37 °C.
Expression of tyrosinase in B16-4A5 and HMV-II cells: Crude tyrosinase was prepared based on the method of Ohguchi et al. [16]. Tyrosinase expression was confirmed by Western blotting [17]. See Supporting Information for detailed protocols.
Supporting information
Detailed information on materials and methods are available as Supporting Information.
Acknowledgements
We thank Dr. Junpei Mutoh of Kyushu University of Health and Welfare for help with Western blotting, and Ms. Misa Uchiyamada and Ms. Kayo Sonoda for their excellent technical assistance.
Conflict of Interest
The authors declare no conflict of interest.
-
References
- 1 Tomira Y, Maeda K, Tagami H. Mechanisms for hyperpigmentation in postinflammatory pigmentation, urticarial pigmentation and sunburn. Dermatologica 1989; 179 (Suppl. 01) 49-53
- 2 Briganti S, Camera E, Picardo M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment Cell Res 2003; 16: 101-110
- 3 Chang TS. An updated review of tyrosinase inhibitors. Int J Mol Sci 2009; 10: 2440-2475
- 4 Jennifer C, Stephie CM, Abhishri SB, Shalini BU. A review on skin whitening property of plant extracts. Int J Pharm Bio Sci 2012; 3: 332-347
- 5 Maeda K, Fukuda M. In vitro effectiveness of several whitening cosmetic components in human melanocyte. J Soc Cosmet Chem 1991; 42: 361-368
- 6 Nakamura T, Seki S, Matsubara O, Ito S, Kasuga T. Specific incorporation of 4-S-cysteinylphenol into human melanoma cells. J Invest Dermatol 1988; 90: 725-728
- 7 Mitani K, Takano F, Kawabata T, Allam AE, Ota M, Takahashi T, Yahagi N, Sakurada C, Fushiya S, Ohta T. Suppression of melanin synthesis by the phenolic constituents of sappanwood (Caesalpinia sappan). Planta Med 2013; 79: 37-44
- 8 Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T. Inhibitory effects of alpha-arbutin on melanin synthesis in cultured human melanoma cells and a three-dimensional human skin model. Biol Pharm Bull 2004; 27: 510-514
- 9 Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T. Syntheses of arbutin-alpha-glycosides and a comparison of their inhibitory effects with those of alpha-arbutin and arbutin on human tyrosinase. Chem Pharm Bull (Tokyo) 2003; 51: 798-801
- 10 Pomerantz SH. The tyrosine hydroxylase activity of mammalian tyrosinase. J Biol Chem 1966; 241: 161-168
- 11 Hwang JH, Lee BM. Inhibitory effects of plant extracts on tyrosinase, L-DOPA oxidation, and melanin synthesis. J Toxicol Environ Health A 2007; 70: 393-407
- 12 Sato K, Takahashi H, Iraha R, Toriyama M. Down-regulation of tyrosinase expression by acetylsalicylic acid in murine B16 melanoma. Biol Pharm Bull 2008; 31: 33-37
- 13 Kai H, Baba M, Okuyama T. Inhibitory effect of Cucumis sativus on melanin production in melanoma B16 cells by downregulation of tyrosinase expression. Planta Med 2008; 74: 1785-1788
- 14 Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T. Inhibitory effects of α-arbutin on melanin synthesis in cultured human melanoma cells and a three-dimensional human skin model. Biol Pharm Bull 2004; 27: 510-514
- 15 Ha SK, Koketsu M, Lee K, Choi SY, Park JH, Ishihara H, Kim SY. Inhibition of tyrosinase activity by N,N-unsubstituted selenourea derivatives. Biol Pharm Bull 2005; 28: 838-840
- 16 Ohguchi K, Akao Y, Nozawa Y. Involvement of calpain in melanogenesis of mouse B16 melanoma cells. Mol Cell Biochem 2005; 275: 103-107
- 17 Hata K, Hori K, Murata J, Takahashi S. Remodeling of actin cytoskeleton in lupeol-induced B16 2F2 cell differentiation. J Biochem 2005; 138: 467-472
Correspondence
-
References
- 1 Tomira Y, Maeda K, Tagami H. Mechanisms for hyperpigmentation in postinflammatory pigmentation, urticarial pigmentation and sunburn. Dermatologica 1989; 179 (Suppl. 01) 49-53
- 2 Briganti S, Camera E, Picardo M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment Cell Res 2003; 16: 101-110
- 3 Chang TS. An updated review of tyrosinase inhibitors. Int J Mol Sci 2009; 10: 2440-2475
- 4 Jennifer C, Stephie CM, Abhishri SB, Shalini BU. A review on skin whitening property of plant extracts. Int J Pharm Bio Sci 2012; 3: 332-347
- 5 Maeda K, Fukuda M. In vitro effectiveness of several whitening cosmetic components in human melanocyte. J Soc Cosmet Chem 1991; 42: 361-368
- 6 Nakamura T, Seki S, Matsubara O, Ito S, Kasuga T. Specific incorporation of 4-S-cysteinylphenol into human melanoma cells. J Invest Dermatol 1988; 90: 725-728
- 7 Mitani K, Takano F, Kawabata T, Allam AE, Ota M, Takahashi T, Yahagi N, Sakurada C, Fushiya S, Ohta T. Suppression of melanin synthesis by the phenolic constituents of sappanwood (Caesalpinia sappan). Planta Med 2013; 79: 37-44
- 8 Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T. Inhibitory effects of alpha-arbutin on melanin synthesis in cultured human melanoma cells and a three-dimensional human skin model. Biol Pharm Bull 2004; 27: 510-514
- 9 Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T. Syntheses of arbutin-alpha-glycosides and a comparison of their inhibitory effects with those of alpha-arbutin and arbutin on human tyrosinase. Chem Pharm Bull (Tokyo) 2003; 51: 798-801
- 10 Pomerantz SH. The tyrosine hydroxylase activity of mammalian tyrosinase. J Biol Chem 1966; 241: 161-168
- 11 Hwang JH, Lee BM. Inhibitory effects of plant extracts on tyrosinase, L-DOPA oxidation, and melanin synthesis. J Toxicol Environ Health A 2007; 70: 393-407
- 12 Sato K, Takahashi H, Iraha R, Toriyama M. Down-regulation of tyrosinase expression by acetylsalicylic acid in murine B16 melanoma. Biol Pharm Bull 2008; 31: 33-37
- 13 Kai H, Baba M, Okuyama T. Inhibitory effect of Cucumis sativus on melanin production in melanoma B16 cells by downregulation of tyrosinase expression. Planta Med 2008; 74: 1785-1788
- 14 Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T. Inhibitory effects of α-arbutin on melanin synthesis in cultured human melanoma cells and a three-dimensional human skin model. Biol Pharm Bull 2004; 27: 510-514
- 15 Ha SK, Koketsu M, Lee K, Choi SY, Park JH, Ishihara H, Kim SY. Inhibition of tyrosinase activity by N,N-unsubstituted selenourea derivatives. Biol Pharm Bull 2005; 28: 838-840
- 16 Ohguchi K, Akao Y, Nozawa Y. Involvement of calpain in melanogenesis of mouse B16 melanoma cells. Mol Cell Biochem 2005; 275: 103-107
- 17 Hata K, Hori K, Murata J, Takahashi S. Remodeling of actin cytoskeleton in lupeol-induced B16 2F2 cell differentiation. J Biochem 2005; 138: 467-472