

Subscribe to RSS
DOI: 10.1055/s-0041-1724076
BMP-4, TGF-β e Smad3 como moduladores da viabilidade das células do líquido sinovial[*]
Article in several languages: português | EnglishAuthors

Resumo
Objetivo Nosso objetivo foi avaliar a modulação das células do líquido sinovial (SFCs, na sigla em inglês) de pacientes com e sem osteoartrite (OA) por proteína morfogenética óssea 4 (BMP-4), Smad3 e transformador do fator de crescimento β (TGF-β).
Métodos O do líquido sinovial foi coletado de pacientes submetidos a artroscopia ou artroplastia do joelho, e centrifugados para isolar as células do liquido sinovial. As células foram cultivadas por 21 dias e caracterizadas como células-tronco mesenquimais (MSCs, na sigla em inglês) de acordo com os critérios da International Society of Cell Therapy. Em seguida, realizamos um ensaio de brometo de 3-4,5-dimetil-tiazol-2-il-2,5-difeniltetrazólio (MTT) depois de expor células com e sem OA para TGF-β, inibidores de via Smad3 e BMP-4 e para diferentes concentrações de BMP-4.
Resultados A exposição aos inibidores TGF-β, Smad3 e BMP-4 modifica a atividade mitocondrial das SFCs. A atividade das SFCs é modificada por influências sobre o aumento das concentrações de BMP-4, mas não há diferença na atividade celular entre pacientes com e sem OA.
Conclusão TGF-β, Smad3 e BMP-4 modulam a atividade das SFCs de pacientes com e sem OA do joelho.
Palavras-chave
células-tronco mesenquimais - osteoartrite - células do líquido sinovial - caminho TGF-βIntrodução
A terapia celular tem sido estudada devido a sua possível aplicação para o tratamento de lesões de cartilagem articular e osteoartrite (OA).[1] [2] [3] [4] Neste contexto, as células-tronco mesenquimais (MSCs) têm sido comprovadamente úteis na regeneração tecidual e no tratamento de muitas doenças.[3] O líquido sinovial também possui uma população residente de MSCs, que é aumentada em pacientes com OA.[5] Além disso, demonstrou-se que o número de células era proporcional à gravidade da OA do joelho.[6]
O transformador do fator de crescimento β (TGF-β) parece ter papel central no desenvolvimento da OA, seja inicialmente como fator protetor ou participando da patogênese da doença.[7] A superfamília TGF-β é um grupo de polipeptídios secretos, incluindo TGF-β1, 2, 3, ativações, inibinas, proteínas morfogenéticas ósseas (BMPs, na sigla em inglês), e fatores de crescimento e diferenciação.[8] [9] Ela atua em todas as etapas de condrogênese e é o principal gatilho através de precursores mesenquimais.[10] [11] [12] Nos primeiros estágios de diferenciação dos condrócitos, o TGF-β parece ter papel estimulante. No entanto, nos estágios finais, sua ação inibe a diferenciação da terminação do condrócito.[12] [13] AS BMPs estão envolvidas em todas as etapas da condrogênese e são essenciais para a ossificação endocondral. Além disso, muitas BMPs foram detectadas tanto na cartilagem normal quanto na osteoartrítica.[9]
A senescência tem impactos nas vias TGF-β e BPM devido à diminuição da expressão dos receptores e à redução da ativação do Smad3. Isso poderia explicar a estreita relação entre senescência e desenvolvimento de OA.[14] Pacientes com OA têm níveis elevados de BMP-2[15] e BMP-7[16] em plasma e líquido sinovial em comparação com indivíduos saudáveis. Além disso, os níveis de BMP-7 estão correlacionados com a magnitude de progressão radiográfica da OA.[17] No entanto, a expressão de BMP-4, que estimula a síntese da matriz extracelular em condrócitos, é diminuída nos tecidos sinoviais de OA e artrite reumatóide (AR), indicando um desequilíbrio da homeostase articular na doença articular.[18] O objetivo do presente estudo foi avaliar o papel de BMP-4, Smad3 e TGF-β na viabilidade de células do líquido sinovial (SFCs, na sigla em inglês) de pacientes com e sem OA.
Métodos
População e Coleta de Líquido Sinovial
A população do estudo foi composta por pacientes submetidos ao tratamento cirúrgico do joelho em nosso Instituto. Os pacientes elegíveis de ambos os sexos foram divididos em dois grupos: pacientes sem OA do joelho (W/O OA), Kellgren-Lawrence = 0, e pacientes com OA do joelho (W/OA), Kellgren-Lawrence > 2. Foram excluídos pacientes com infecção do joelho, doenças autoimunes ou submetidos a cirurgias anteriores no joelho. A OA de joelho foi classificada de acordo com a classificação de Kellgren e Lawrence usando radiografias padrão em pé.[19] Um mililitro de fluido sinovial foi coletado da articulação do joelho dos pacientes W/O OA no início da artroscopia por aspiração de seringa, logo após a confecção dos portais, e dois mililitros foram coletados da articulação do joelho dos pacientes W/OA por aspiração de seringa após artrotomia. O presente estudo foi realizado de acordo com os princípios da Declaração de Helsinque e foi aprovado pelo Conselho de Ética Institucional (CAAE 08663912.3.0000.5273).
Isolamento e Cultura de Células do Líquido Sinovial
Os espécimes de líquido sinovial foram primeiro centrifugados a 16,6 g por 5 minutos em temperatura ambiente, e depois a 1.600 g por 5 minutos a 4°C. O sobrenadante foi armazenado em partes de 1 mL a - 80°C para análise posterior. O pellet da primeira centrifugação foi suspenso no Meio de Dulbecco Modificado por Iscove (IMDM, Sigma, Saint Louis, Missouri) complementado com 10% de soro bovino fetal (FBS, Gibco, Thermo Fisher, Waltham, MA, EUA). As células foram semeadas em 5 a 7 × 105 células por frascos de cultura T-75 cm2. Após 24 horas, as células foram lavadas uma vez com meio livre de soro e o meio foi trocado. As culturas celulares foram mantidas a 37°C em uma atmosfera umidificada contendo 5% de CO2. O meio era trocado a cada 2 dias. As células foram colhidas e rebanhadas para expansão em 80% da confluência até a 3ª passagem. A identidade das SFCs foi avaliada de acordo com o protocolo definido pela International Society of Cell Therapy[20] e descrita em um estudo anterior.[21]
Atividade Mitocondrial SFC por MTT
Para avaliar a atividade mitocondrial do SFC exposta a diferentes concentrações de BMP-4, foi realizado um ensaio de atividade mitocondrial por ensaio de brometo de 3-4,5-dimetil-tiazol-2-il-2,5-difeniltetrazólio (MTT) (4-[4,5-dimetilthiazol-2-yl]-2,5-diphenyltetrazolium brometo). Em uma placa de 96 poços, 5 × 103 SFC de pacientes W/O OA (n = 3) e W/OA (n = 3) foram semeados em 200µL de meio/poço e expostos a concentrações progressivas de BMP-4 (0,1 nM, 0,5 nM, 2 nM e 10 nM) e mantido a 37°C em uma atmosfera umidificada contendo 5% de CO2.
O controle da atividade mitocondrial do SFC dos pacientes W/O OA (n = 3) e W/OA (n = 3) foi avaliado pelo bloqueio das vias BMP-4, TGF-β e Smad3. Como tal, o SFC foi cultivado na presença dos inibidores específicos dos alvos Smad3 (Smad3 SiS3 566405, Millipore, Burlington, Massachusetts, EUA), BMP-4 (inibidor de BMP II DMH1 203646, Merckmillipore, Burlington, Massachusetts, EUA) e TGF-β (TGF-β1 R1 quinase 616451, Merckmillipore, Burlington, Massachusetts, EUA) a 10 nM.
Após a adesão celular, o meio de cultura foi substituído pelo meio de Iscove (Sigma-Aldrich, San Luis, Missouri, EUA) suplementado com 1% de soro bovino fetal (Sigma-Aldrich, San Luis, Missouri, EUA) contendo os inibidores específicos para cada tratamento. O SFC foi tratado com 100 μL das respectivas soluções de controle e experimentais em 37°C em uma atmosfera umidificada contendo 5% de CO2 para 24, 48 e 72 horas. Em seguida, o SFC foi lavado 2 vezes com 200 μL PBS 1x estéril a 37°C e 100 μL da solução MTT (0,5mg/mL) foi adicionado a cada poço. A placa foi mantida a 37°C em uma atmosfera umidificada contendo 5% de CO2 durante 4 horas.
Após esse período, a solução MTT foi removida e 100 μL dimetilsulfóxido (DMSO) foi adicionada. A placa foi agitada por 20 minutos antes da análise espectrofotométrica em comprimento de onda de 570 nM usando o sistema de detecção Glomaxmulti (Glomaxmulti, Sistema de detecção, Promega, Madison, Wisconsin, EUA). A análise da atividade mitocondrial do SFC foi medida após 24, 48 e 72 horas de exposição a cada inibidor e controle específico. O controle da atividade mitocondrial foi determinado pelo MTT tanto por poços contendo apenas meio com BMP4 sem células (controle/fator) quanto por poços contendo apenas meio e células, mas sem BMP4.
Análise Estatística
Os dados MTT foram exportados do sistema de detecção Glomaxmulti (Glomaxmulti, Sistema de Detecção, Promega, Madison, Wisconsin, EUA) para uma planilha de Microsoft Excel (Microsoft Corporation, Redmond, WA, EUA), analisada no GraphPad Prism para Windows 6.0 (San Diego, California, EUA) e apresentada como ± desvio-padrão médio. Para comparar grupos, foi utilizada a análise de variância (ANOVA) bidirecional, seguida pelo teste t de Student não pareado unicaudal. Os dados foram considerados significativos quando p < 0,05.
Resultados
BMP-4 modula a viabilidade de SFC
A atividade mitocondrial por MTT não aumentou as diferenças entre as amostras de pacientes com e sem OA de joelho quando analisadas em cada situação experimental, independentemente da concentração de BMP-4 (p > 0,05). Quando analisados os dados em relação às concentrações crescentes de BMP-4, verifica-se que a atividade mitocondrial das SFCs de pacientes com OA aumentou progressivamente de acordo com a exposição a maiores concentrações de BMP-4 (0,1 nM 264,4 ± 11,94 versus 0,5 nM 562,8 ± 26,17, p < 0.0001; 0,5 nM 562,8 ± 26,17 versus 2 nM 1157 ± 40,45, p < 0,0001), diminuindo na exposição à concentração de 10 nM (2 nM 1157 ± 40,45 versus 10 nM 124,9 ± 14,62, p < 0,0001), mostrando toxicidade. O mesmo foi observado para a atividade mitocondrial das SFCs de pacientes sem OA (0,1 nM 141,7 ± 20,83 versus 0,5 nM 496,3 ± 62,44, p < 0,0001; 0,5 nM 496,3 ± 62,44 versus 2 nM 1134 ± 69,22, p < 0,0001; 2 nM 1134 ± 69,22 versus 10 nM 168,2 ± 20,33, p < 0,0001) ([Figura 1]).

Inibidores TGF-β, Smad3 e BMP-4 controlam atividade mitocondrial das SFCs
No passo seguinte, medimos a atividade mitocondrial das SFCs após a exposição aos inibidores TGF-β, Smad3 e BMP-4 por ensaio MTT [4-(4,5-dimetilthiazol-2-yl)-2,5-difeniltetrazolium]. As SFCs de pacientes com e sem OA de joelho foram expostas a inibidores TGF-β (TGF-β1 R1 quinase 616451), Smad3 (Smad3 SiS3 566405) e BMP-4 (inibidor de BMP II DMH1 203646) na concentração de 10 nM. Em seguida, a atividade mitocondrial foi medida após 24, 48 e 72 horas.
A análise ([Figura 2]) evidencia uma diminuição das SFCs sem atividade de OA após 24 horas na presença do inibidor de TGFβ (W/O OA 202,1 ± 6,09 versus W/O OA + inibidor de β 175,7 ± 9,56; n = 3/grupo; p = 0,016) e aumento da atividade das SFCs após 48 horas (W/OA 190 ± 6,5 versus W/OA + TGF-inibidor de β 242 ± 2,5; n = 3/grupo; p < 0,0001). Não foram encontradas diferenças após 72 horas (W/O OA 134,3 ± 5,1 versus W/O OA + inibidor TGF-β 135 ± 3,01; n = 3/grupo; p > 0,05). Em relação às SFCs de pacientes com OA, a análise mostrou aumento da atividade das SFCs somente após 48 horas na presença do inibidor de TGF-β (W/OA 215 ± 2,14 versus W/OA + inibidor TGF-β 229,3 ± 4,16; n = 3/grupo; p = 0,006). Não foram encontradas diferenças após 24 horas (W/OA 175,2 ± 17,72 versus W/OA + inibidor TGF-β 173,9 ± 10,34; n = 3/grupo; p > 0,05) e 72 horas (W/OA 140,7 ± 3,2 versus W/OA + inibidor TGF-β 144 ± 3,6; n = 3/grupo; p > 0,05).

A atividade das SFCs sem OA reduziu após 24 horas na presença do inibidor Smad3 (W/O OA 202.1 ± 6,09 versus W/O OA + inibidor Smad3 189,7 ± 3,39; n = 3/grupo; p = 0,046), mas aumentou após 48 horas (W/O OA 190 ± 6,5 versus W/OA + inibidor Smad3 224 ± 4,04; n = 3/grupo; p = 0,0002) e 72 horas (W/O OA 134,3 ± 5,17 versus W/O OA + inibidor Smad3 158 ± 6,5; n = 3/grupo; p = 0,006) na presença do inibidor Smad3. Em relação às SFCs de pacientes com OA, a análise evidencia a redução da atividade somente após 72 horas na presença do inibidor Smad3 (W/OA 162 ± 3,7 versus inibidor W/OA + Smad3 151 ± 2; n = 3/grupo; p = 0,0102). Não foram encontradas diferenças após 24 horas (W/OA 180 ± 7,4 versus W/OA + inibidor Smad3 318,2 ± 5,99; n = 3/grupo; p > 0,05) e 48 horas (W/OA 212,3 ± 6,5 versus W/OA + inibidor Smad3 206,7 ± 5,8; n = 3/grupo; p > 0,05) ([Figura 3]).
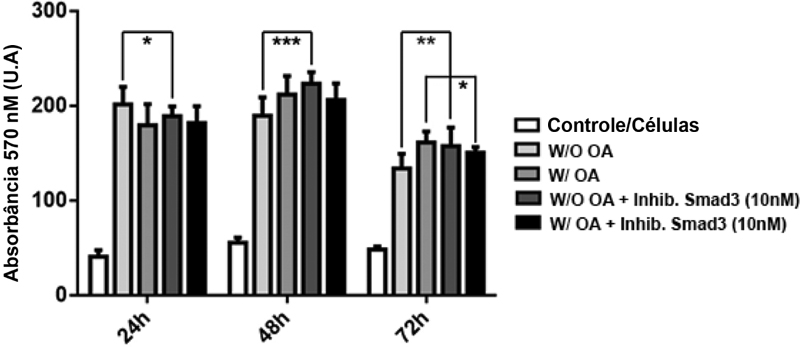
A atividade das SFCs sem OA aumentou após 48 horas (W/O OA 134,3 ± 5,17 versus W/O + inibidor BMP4 177,3 ± 14,45; n = 3/grupo; p = 0,0064) e 72 horas (W/O 173,3 ± 1,83 versus W/O + inibidor BMP4 243,7 ± 13,15; n = 3/grupo; p < 0,0001) na presença do inibidor BMP-4. As SFCs com atividade OA também aumentaram após 48 horas (W/OA 159 ± 3,21 versus W/OA + inibidor BMP4 218 ± 8,94; n = 3/grupo; p < 0,0001) e 72 horas (W/OA 158 ± 2,29 versus inibidor BMP4 240,3 ± 0,83; n = 3/grupo; p < 0,0001) na presença do inibidor BMP-4. Não houve diferenças na atividade das SFCs sem OA (W/O 179,9 ± 16,27 versus W/O + inibidor BMP4 156,8 ± 16,27; n = 3/grupo; p > 0,05) e com OA (W/OA 140,1 ± 14,59 versus inibidor BMP4 172,9 ± 21,15; n = 3/grupo; p > 0,05) após 24 horas na presença do inibidor BMP-4 ([Figura 4]).

Discussão
Documentamos anteriormente que o líquido sinovial osteoartrítico modula a viabilidade das SFCs in vitro.[21] Por isso, decidimos investigar se a atividade metabólica das SFCs poderia ser modulada por BMP-4, Smad3 e TGF-β, devido à importância dessas vias na condrogênese.
O BMP-4 estimula a síntese de aggrecan e colágeno tipo II e suprime a hipertrofia condrogênica, sugerindo que pode ser um agente promissor para a indução de reparação de cartilagem. A modulação do BMP pode ser útil em doenças articulares crônicas, pois pode ajudar no equilíbrio entre destruição articular e reparação.[22] Para elucidar como as SFCs se comportam na presença de BMP-4 exógeno, no presente estudo, mostramos que a atividade metabólica das SFCs aumenta após a exposição às concentrações progressivas de BMP-4 de 0,1 a 2 nM, sugerindo que as SFCs estão respondendo à indução de BMP-4 a fim de promover a reparação, talvez por estímulo ao comprometimento condrogênico, diminuindo a 10 nM, que parece ser uma concentração na qual o sistema já está saturado, não influenciando mais no destino das SFCs. Além disso, não houve diferenças nessa atividade ao comparar as SFCs de pacientes com e sem OA, indicando que, apesar da influência inflamatória da OA nas SFCs, elas ainda mantêm a capacidade de responder à sinalização BMP-4.
Por outro lado, observou-se também que a atividade mitocondrial aumentou após 48 e 72 horas na presença do inibidor BMP-4. É amplamente conhecido que o BMP-4 modula a condrogênese e a ossificação endocondral,[23] assim como participa da sinalização para proliferação de condrócitos e hipertrofia à apoptose e angiogênese que permite a migração e ossificação dos osteoblastos. O bloqueio do receptor BMP-4 levou à diminuição da expressão da sinalização BMP-4, e elevou a atividade metabólica das SFCs, sugerindo que o passo condrogênico da diferenciação das SFCs permaneceu no estágio proliferativo, o que poderia explicar o aumento da atividade metabólica das SFCs. Apesar da importância do BMP-4, não encontramos estudos sobre sua influência no comportamento das SFCs. Outros estudos, no entanto, verificaram que o pré-tratamento das SFCs com TNF-β[24] e IL-1β[25] promoveu sua proliferação. Nossos resultados indicados no tratamento das SFCs tanto com inibidor BMP-4 ou BMP-4 podem influenciar a proliferação de SFCs, dependendo da magnitude dos estímulos.
A presença de TGF-β no líquido sinovial de pacientes com OA tem um papel importante no recrutamento de MSC. Além disso, o líquido sinovial de pacientes com OA pode estimular a expansão do MSC na cultura de células sinoviais de pacientes com OA, através da estimulação da migração celular.[26] Outro estudo mostrou que a expressão contínua de TGF-β1 sobre o MSC sinovial estimulou sua proliferação e potencial condrogênico.[27]
Durante a condrogênese, a TGF-β é importante para a diferenciação de condrócitos em estágios iniciais e, por fim, na indução condrogênica, inibe a diferenciação de condrócitos terminais.[12] Além disso, a TGF-β promove o crescimento, a manutenção e a reparação da cartilagem articular por meio de uma boa regulação de sua via de sinalização, que visa um conjunto de fatores de transcrição e crescimento.[28] Fenômenos celulares regulados incluem proliferação, migração, inflamação, carcinogênese, expressão de matriz extracelular, síntese proteica e degradação, até funções do sistema imunológico. Muitos autores afirmaram que a sinalização de vias TGF-β tem um papel central no desenvolvimento da OA.[7] [12] [14] [29] Como observamos no presente estudo, o bloqueio de TGF-β promoveu uma mudança dinâmica na atividade mitocondrial. Nas primeiras 24 horas, foi observada uma redução em pacientes sem OA na presença do inibidor TGF-β. A inibição do receptor Smad3 diminuiu a atividade apenas nas SFCs de pacientes sem OA. Surpreendentemente, após 48 horas de inibição, a atividade aumentou nas SFCs de pacientes com e sem OA na presença do inibidor TGF-β. A inibição do Smad3 promoveu um aumento na atividade das SFCs apenas de pacientes sem OA neste mesmo período. Após 72 horas de exposição ao inibidor TGF-β, a atividade foi semelhante em ambos os grupos de SFCs, mas aumentou na presença do inibidor Smad3 no caso de SFCs de pacientes sem OA e diminuiu nos com OA.
Conclusão
Em conclusão, não há diferenças na atividade mitocondrial das SFCs de pacientes com e sem OA, independentemente da concentração BMP-4 à qual foram expostas. Além disso, a atividade das SFCs de pacientes com e sem OA é modulada pelos inibidores de TGF-β, Smad 3 e BMP-4.
Conflitos de Interesse
Os autores declaram não haver conflitos de interesse.
Agradecimentos
Os autores agradecem ao Conselho Nacional de Desenvolvimento Científico e Tecnológico, à Fundação de Apoio à Pesquisa do Estado do Rio de Janeiro e ao Ministério da Saúde pelo apoio.
Contribuições autorais
de Sousa EB Conceito e projeto, fornecimento de material de estudo ou pacientes, coleta e montagem de dados, análise e interpretação de dados, redação de manuscritos, aprovação final do manuscrito.
Moura Neto V.: Análise e interpretação de dados, aprovação final do manuscrito.
Aguiar D. P.: Conceito e projeto, coleta e montagem de dados, análise e interpretação de dados, redação de manuscritos, aprovação final do manuscrito.
Todos os autores leram e aprovaram o manuscrito final.
* Trabalho desenvolvido na Divisão de Ensino e Pesquisa, Instituto Nacional de Ortopedia e Traumatologia Jamil Haddad, Rio de Janeiro, RJ, Brasil.
-
Referências
- 1 Gupta PK, Das AK, Chullikana A, Majumdar AS. Mesenchymal stem cells for cartilage repair in osteoarthritis. Stem Cell Res Ther 2012; 3 (04) 25
- 2 Johnson K, Zhu S, Tremblay MS. et al. A stem cell-based approach to cartilage repair. Science 2012; 336 (6082): 717-721
- 3 Barry F, Murphy M. Mesenchymal stem cells in joint disease and repair. Nat Rev Rheumatol 2013; 9 (10) 584-594
- 4 Pers YM, Ruiz M, Noël D, Jorgensen C. Mesenchymal stem cells for the management of inflammation in osteoarthritis: state of the art and perspectives. Osteoarthritis Cartilage 2015; 23 (11) 2027-2035
- 5 Jones EA, Crawford A, English A. et al. Synovial fluid mesenchymal stem cells in health and early osteoarthritis: detection and functional evaluation at the single-cell level. Arthritis Rheum 2008; 58 (06) 1731-1740
- 6 Sekiya I, Ojima M, Suzuki S. et al. Human mesenchymal stem cells in synovial fluid increase in the knee with degenerated cartilage and osteoarthritis. J Orthop Res 2012; 30 (06) 943-949
- 7 van der Kraan PM, Goumans MJ, Blaney Davidson E, ten Dijke P. Age-dependent alteration of TGF-β signalling in osteoarthritis. Cell Tissue Res 2012; 347 (01) 257-265
- 8 Keller B, Yang T, Chen Y. et al. Interaction of TGFβ and BMP signaling pathways during chondrogenesis. PLoS One 2011; 6 (01) e16421
- 9 Mariani E, Pulsatelli L, Facchini A. Signaling pathways in cartilage repair. Int J Mol Sci 2014; 15 (05) 8667-8698
- 10 Leonard CM, Fuld HM, Frenz DA, Downie SA, Massagué J, Newman SA. Role of transforming growth factor-beta in chondrogenic pattern formation in the embryonic limb: stimulation of mesenchymal condensation and fibronectin gene expression by exogenenous TGF-beta and evidence for endogenous TGF-beta-like activity. Dev Biol 1991; 145 (01) 99-109
- 11 Song JJ, Aswad R, Kanaan RA. et al. Connective tissue growth factor (CTGF) acts as a downstream mediator of TGF-beta1 to induce mesenchymal cell condensation. J Cell Physiol 2007; 210 (02) 398-410
- 12 van der Kraan PM, Blaney Davidson EN, Blom A, van den Berg WB. TGF-beta signaling in chondrocyte terminal differentiation and osteoarthritis: modulation and integration of signaling pathways through receptor-Smads. Osteoarthritis Cartilage 2009; 17 (12) 1539-1545
- 13 Ballock RT, Heydemann A, Wakefield LM, Flanders KC, Roberts AB, Sporn MB. TGF-beta 1 prevents hypertrophy of epiphyseal chondrocytes: regulation of gene expression for cartilage matrix proteins and metalloproteases. Dev Biol 1993; 158 (02) 414-429
- 14 van Caam A, Madej W, Thijssen E. et al. Expression of TGFβ-family signalling components in ageing cartilage: age-related loss of TGFβ and BMP receptors. Osteoarthritis Cartilage 2016; 24 (07) 1235-1245
- 15 Liu Y, Hou R, Yin R, Yin W. Correlation of bone morphogenetic protein-2 levels in serum and synovial fluid with disease severity of knee osteoarthritis. Med Sci Monit 2015; 21: 363-370
- 16 Honsawek S, Chayanupatkul M, Tanavalee A. et al. Relationship of plasma and synovial fluid BMP-7 with disease severity in knee osteoarthritis patients: a pilot study. Int Orthop 2009; 33 (04) 1171-1175
- 17 Caron MM, Emans PJ, Cremers A. et al. Hypertrophic differentiation during chondrogenic differentiation of progenitor cells is stimulated by BMP-2 but suppressed by BMP-7. Osteoarthritis Cartilage 2013; 21 (04) 604-613
- 18 Bramlage CP, Häupl T, Kaps C. et al. Decrease in expression of bone morphogenetic proteins 4 and 5 in synovial tissue of patients with osteoarthritis and rheumatoid arthritis. Arthritis Res Ther 2006; 8 (03) R58
- 19 Kellgren JH, Lawrence JS. Radiological assessment of osteo-arthrosis. Ann Rheum Dis 1957; 16 (04) 494-502
- 20 Dominici M, Le Blanc K, Mueller I. et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006; 8 (04) 315-317
- 21 de Sousa EB, Dos Santos Junior GC, Aguiar RP. et al. Osteoarthritic Synovial Fluid Modulates Cell Phenotype and Metabolic Behavior In Vitro . Stem Cells Int 2019; 2019: 8169172
- 22 Miljkovic ND, Cooper GM, Marra KG. Chondrogenesis, bone morphogenetic protein-4 and mesenchymal stem cells. Osteoarthritis Cartilage 2008; 16 (10) 1121-1130
- 23 Carreira AC, Alves GG, Zambuzzi WF, Sogayar MC, Granjeiro JM. Bone Morphogenetic Proteins: structure, biological function and therapeutic applications. Arch Biochem Biophys 2014; 561: 64-73
- 24 Shioda M, Muneta T, Tsuji K. et al. TNFα promotes proliferation of human synovial MSCs while maintaining chondrogenic potential. PLoS One 2017; 12 (05) e0177771
- 25 Matsumura E, Tsuji K, Komori K, Koga H, Sekiya I, Muneta T. Pretreatment with IL-1β enhances proliferation and chondrogenic potential of synovium-derived mesenchymal stem cells. Cytotherapy 2017; 19 (02) 181-193
- 26 Zhang S, Muneta T, Morito T, Mochizuki T, Sekiya I. Autologous synovial fluid enhances migration of mesenchymal stem cells from synovium of osteoarthritis patients in tissue culture system. J Orthop Res 2008; 26 (10) 1413-1418
- 27 Kim YI, Ryu JS, Yeo JE. et al. Overexpression of TGF-β1 enhances chondrogenic differentiation and proliferation of human synovium-derived stem cells. Biochem Biophys Res Commun 2014; 450 (04) 1593-1599
- 28 Finnson KW, Chi Y, Bou-Gharios G, Leask A, Philip A. TGF-b signaling in cartilage homeostasis and osteoarthritis. Front Biosci (Schol Ed) 2012; 4: 251-268
- 29 Plaas A, Velasco J, Gorski DJ. et al. The relationship between fibrogenic TGFβ1 signaling in the joint and cartilage degradation in post-injury osteoarthritis. Osteoarthritis Cartilage 2011; 19 (09) 1081-1090
Endereço para correspondência
Publication History
Received: 31 May 2020
Accepted: 16 September 2020
Article published online:
31 March 2021
© 2021. Sociedade Brasileira de Ortopedia e Traumatologia. This is an open access article published by Thieme under the terms of the Creative Commons Attribution-NonDerivative-NonCommercial License, permitting copying and reproduction so long as the original work is given appropriate credit. Contents may not be used for commecial purposes, or adapted, remixed, transformed or built upon. (https://creativecommons.org/licenses/by-nc-nd/4.0/)
Thieme Revinter Publicações Ltda.
Rua do Matoso 170, Rio de Janeiro, RJ, CEP 20270-135, Brazil
-
Referências
- 1 Gupta PK, Das AK, Chullikana A, Majumdar AS. Mesenchymal stem cells for cartilage repair in osteoarthritis. Stem Cell Res Ther 2012; 3 (04) 25
- 2 Johnson K, Zhu S, Tremblay MS. et al. A stem cell-based approach to cartilage repair. Science 2012; 336 (6082): 717-721
- 3 Barry F, Murphy M. Mesenchymal stem cells in joint disease and repair. Nat Rev Rheumatol 2013; 9 (10) 584-594
- 4 Pers YM, Ruiz M, Noël D, Jorgensen C. Mesenchymal stem cells for the management of inflammation in osteoarthritis: state of the art and perspectives. Osteoarthritis Cartilage 2015; 23 (11) 2027-2035
- 5 Jones EA, Crawford A, English A. et al. Synovial fluid mesenchymal stem cells in health and early osteoarthritis: detection and functional evaluation at the single-cell level. Arthritis Rheum 2008; 58 (06) 1731-1740
- 6 Sekiya I, Ojima M, Suzuki S. et al. Human mesenchymal stem cells in synovial fluid increase in the knee with degenerated cartilage and osteoarthritis. J Orthop Res 2012; 30 (06) 943-949
- 7 van der Kraan PM, Goumans MJ, Blaney Davidson E, ten Dijke P. Age-dependent alteration of TGF-β signalling in osteoarthritis. Cell Tissue Res 2012; 347 (01) 257-265
- 8 Keller B, Yang T, Chen Y. et al. Interaction of TGFβ and BMP signaling pathways during chondrogenesis. PLoS One 2011; 6 (01) e16421
- 9 Mariani E, Pulsatelli L, Facchini A. Signaling pathways in cartilage repair. Int J Mol Sci 2014; 15 (05) 8667-8698
- 10 Leonard CM, Fuld HM, Frenz DA, Downie SA, Massagué J, Newman SA. Role of transforming growth factor-beta in chondrogenic pattern formation in the embryonic limb: stimulation of mesenchymal condensation and fibronectin gene expression by exogenenous TGF-beta and evidence for endogenous TGF-beta-like activity. Dev Biol 1991; 145 (01) 99-109
- 11 Song JJ, Aswad R, Kanaan RA. et al. Connective tissue growth factor (CTGF) acts as a downstream mediator of TGF-beta1 to induce mesenchymal cell condensation. J Cell Physiol 2007; 210 (02) 398-410
- 12 van der Kraan PM, Blaney Davidson EN, Blom A, van den Berg WB. TGF-beta signaling in chondrocyte terminal differentiation and osteoarthritis: modulation and integration of signaling pathways through receptor-Smads. Osteoarthritis Cartilage 2009; 17 (12) 1539-1545
- 13 Ballock RT, Heydemann A, Wakefield LM, Flanders KC, Roberts AB, Sporn MB. TGF-beta 1 prevents hypertrophy of epiphyseal chondrocytes: regulation of gene expression for cartilage matrix proteins and metalloproteases. Dev Biol 1993; 158 (02) 414-429
- 14 van Caam A, Madej W, Thijssen E. et al. Expression of TGFβ-family signalling components in ageing cartilage: age-related loss of TGFβ and BMP receptors. Osteoarthritis Cartilage 2016; 24 (07) 1235-1245
- 15 Liu Y, Hou R, Yin R, Yin W. Correlation of bone morphogenetic protein-2 levels in serum and synovial fluid with disease severity of knee osteoarthritis. Med Sci Monit 2015; 21: 363-370
- 16 Honsawek S, Chayanupatkul M, Tanavalee A. et al. Relationship of plasma and synovial fluid BMP-7 with disease severity in knee osteoarthritis patients: a pilot study. Int Orthop 2009; 33 (04) 1171-1175
- 17 Caron MM, Emans PJ, Cremers A. et al. Hypertrophic differentiation during chondrogenic differentiation of progenitor cells is stimulated by BMP-2 but suppressed by BMP-7. Osteoarthritis Cartilage 2013; 21 (04) 604-613
- 18 Bramlage CP, Häupl T, Kaps C. et al. Decrease in expression of bone morphogenetic proteins 4 and 5 in synovial tissue of patients with osteoarthritis and rheumatoid arthritis. Arthritis Res Ther 2006; 8 (03) R58
- 19 Kellgren JH, Lawrence JS. Radiological assessment of osteo-arthrosis. Ann Rheum Dis 1957; 16 (04) 494-502
- 20 Dominici M, Le Blanc K, Mueller I. et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006; 8 (04) 315-317
- 21 de Sousa EB, Dos Santos Junior GC, Aguiar RP. et al. Osteoarthritic Synovial Fluid Modulates Cell Phenotype and Metabolic Behavior In Vitro . Stem Cells Int 2019; 2019: 8169172
- 22 Miljkovic ND, Cooper GM, Marra KG. Chondrogenesis, bone morphogenetic protein-4 and mesenchymal stem cells. Osteoarthritis Cartilage 2008; 16 (10) 1121-1130
- 23 Carreira AC, Alves GG, Zambuzzi WF, Sogayar MC, Granjeiro JM. Bone Morphogenetic Proteins: structure, biological function and therapeutic applications. Arch Biochem Biophys 2014; 561: 64-73
- 24 Shioda M, Muneta T, Tsuji K. et al. TNFα promotes proliferation of human synovial MSCs while maintaining chondrogenic potential. PLoS One 2017; 12 (05) e0177771
- 25 Matsumura E, Tsuji K, Komori K, Koga H, Sekiya I, Muneta T. Pretreatment with IL-1β enhances proliferation and chondrogenic potential of synovium-derived mesenchymal stem cells. Cytotherapy 2017; 19 (02) 181-193
- 26 Zhang S, Muneta T, Morito T, Mochizuki T, Sekiya I. Autologous synovial fluid enhances migration of mesenchymal stem cells from synovium of osteoarthritis patients in tissue culture system. J Orthop Res 2008; 26 (10) 1413-1418
- 27 Kim YI, Ryu JS, Yeo JE. et al. Overexpression of TGF-β1 enhances chondrogenic differentiation and proliferation of human synovium-derived stem cells. Biochem Biophys Res Commun 2014; 450 (04) 1593-1599
- 28 Finnson KW, Chi Y, Bou-Gharios G, Leask A, Philip A. TGF-b signaling in cartilage homeostasis and osteoarthritis. Front Biosci (Schol Ed) 2012; 4: 251-268
- 29 Plaas A, Velasco J, Gorski DJ. et al. The relationship between fibrogenic TGFβ1 signaling in the joint and cartilage degradation in post-injury osteoarthritis. Osteoarthritis Cartilage 2011; 19 (09) 1081-1090



