
RSS-Feed abonnieren
DOI: 10.1055/s-0042-107801
Inhibitory Effects of Pisiferic Acid on Angiogenesis and Lymphangiogenesis
Correspondence
Publikationsverlauf
received 24. August 2015
revised 17. November 2015
accepted 10. April 2016
Publikationsdatum:
19. August 2016 (online)
Abstract
Pisiferic acid, a diterpene isolated from Chamaecyparis pisifera, has a chemical structure similar to carnosic acid, which has antioxidant activity and exerts various biological effects such as antibiotic and antiangiogenic activity. However, the effect of pisiferic acid on angiogenesis, which is involved in various diseases including cancer and diabetic retinopathy, has not been elucidated. We found that pisiferic acid inhibited angiogenesis and suppressed human umbilical vein endothelial cell tube formation and proliferation but had no effect on human umbilical vein endothelial cell chemotaxis. Furthermore, in lymphangiogenesis assays, pisiferic acid inhibited lymphangiogenesis and lymphatic endothelial cell tube formation and proliferation. Together, these results suggest pisiferic acid to be a useful natural compound, such as carnosic acid, with beneficial effects on angiogenesis- and lymphangiogenesis-related diseases.
#
Key words
Chamaecyparis pisifera - Cupressaceae - pisiferic acid - angiogenesis - lymphangiogenesis - endothelial cellsAbbreviations
PA is an abundant natural product found in the leaves of the Japanese tree Sawara [Chamaecyparis pisifera (Siebold & Zucc.) Endl.; Cupressaceae] ([Fig. 1 A]) [1], and has been reported to have various biological activities including DNA polymerase inhibition [2], chitin synthase inhibition [3], and antibiotic activity [4]. It also affects cell signaling pathways in HL60 cells and yeast [5], [6]. PA has been used for the synthesis of its derivative CA ([Fig. 1 B]) [7]. CA has been extensively studied and reported to have various biological activities such as antioxidant activity, promotion of neurite outgrowth, and a protective effect on the nerve system [8], [9], [10]. Recently, we have reported that CA is an inhibitor of angiogenesis [11]. Angiogenesis refers to the formation of new blood vessels and is involved in various diseases such as cancer, diabetic retinopathy, and rheumatoid arthritis [12]. Thus, natural products that inhibit angiogenesis have drawn attention for the development of angiogenesis inhibitors. While CA has been known to inhibit angiogenesis, the effect of PA on angiogenesis has not been reported thus far. In this study, we examined the effect of PA on angiogenesis, and, furthermore, on lymphangiogenesis, which is also involved in inflammation and various diseases [13].

In an ex vivo angiogenesis assay using the rat aortic ring, PA suppressed the outgrowth of microvessels that grew in a week, as shown in [Fig. 2]. At 50 µM, PA completely inhibited angiogenesis, whereas CA suppressed angiogenesis by about 40 % at the same concentration. However, PA did not show a suppressive effect at 25 µM, but CA significantly suppressed angiogenesis at this concentration [11].

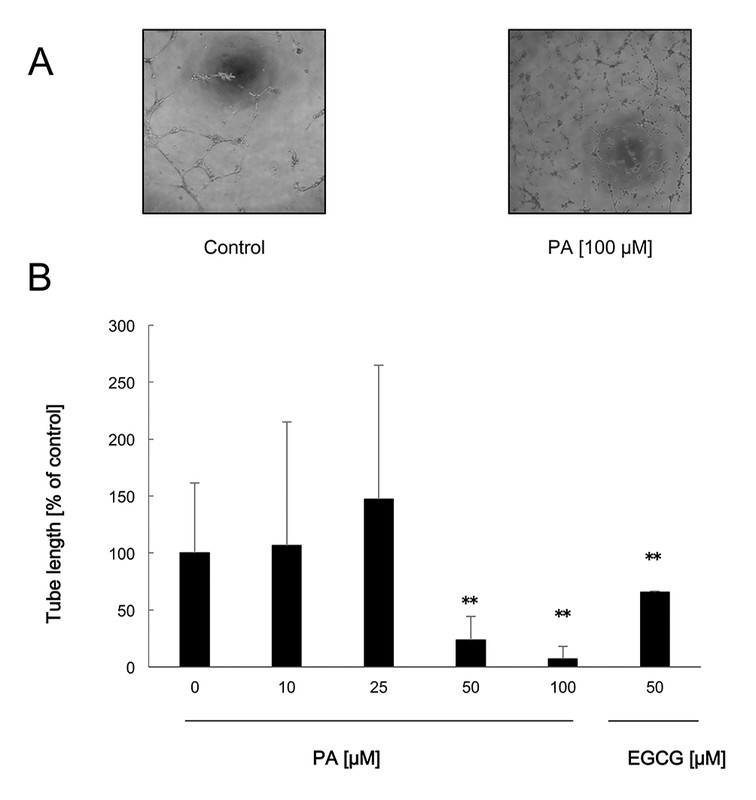
To determine how PA inhibited angiogenesis, we examined its effect in some models using HUVECs. PA significantly suppressed HUVEC tube formation and proliferation at 50 µM and 25 µM, respectively ([Figs. 3] and [4]), but had no effect on HUVEC migration induced by VEGF ([Fig. 5]). Thus, PA exerts antiangiogenic activity by suppressing endothelial cell capillary formation and proliferation. CA exerted suppressive effects in all three HUVEC assays as previously reported [11]. These results suggest that the inhibitory mechanisms by which both compounds exert antiangiogenic activity are different, and that the difference could be attributed to their structural difference. In the HUVEC migration assay, VEGF was used to induce HUVEC migration, but in HUVEC tube formation and proliferation assays, the used medium contained bFGF, and not VEGF, suggesting that PA may not affect the VEGF receptor-mediated cell signaling pathway. Recently, the antiangiogenic activity of fucoxanthin and siphonaxanthin, both natural compounds, has been reported [14], [15]. Both carotenoids did not affect VEGF-induced HUVEC migration, and we demonstrated that both carotenoids downregulated the FGF-2 (bFGF) signaling pathway [16]. Thus, PA might exert its inhibitory effect on angiogenesis in a similar manner. Other derivatives of PA might have unique effects on endothelial cells and angiogenesis.



Lymphangiogenesis is also an important target for preventing various diseases such as cancer and inflammatory conditions. To clarify whether PA affects lymphangiogenesis, we examined its effect in an ex vivo lymphangiogenesis model [17]. Since the method was slightly modified, immunohistochemical analysis was performed to confirm the induction of lymphangiogenesis in the model. The morphology of capillaries was the same as that reported previously ([Fig. 6]), and the anti-LYVE-1 antibody reacted with the capillaries (data not shown), confirming the occurrence of lymphangiogenesis. PA completely inhibited lymphangiogenesis at 100 µM and strongly inhibited it at 50 µM in the model ([Fig. 6]). Since PA suppressed HUVEC tube formation and proliferation, we examined its effect on LEC tube formation and proliferation. PA exerted inhibitory effects in both assays ([Figs. 7] and [8]), suggesting that PA exerts inhibitory effects targeting the same molecules on both endothelial cells for angiogenesis and lymphangiogenesis. Recently, it has been demonstrated that FGF-2 (bFGF) and VEGF-C, an angiogenesis and lymphangiogenesis enhancing growth factor, respectively, collaboratively stimulate tumor growth, angiogenesis, lymphangiogenesis, and metastasis [18]. Thus, natural products suppressing both angiogenesis and lymphangiogenesis or the bFGF pathway would be useful as anticancer agents. Although elucidation of the mechanism by which PA exerts antilymphangiogenic activity is necessary, this study suggests that PA might be beneficial in suppressing both angiogenesis- and lymphangiogenesis-related diseases.


In this study, we found that PA suppresses both angiogenesis and lymphangiogenesis. We previously reported the antiangiogenic effect of CA, whose chemical structure is very similar to PA. The inhibitory effects of PA on angiogenesis are similar to that of CA, but its effect on HUVECs is different. CA suppresses HUVEC tube formation, proliferation, and migration, whereas PA has no effect on HUVEC migration. This difference in biological activity could be attributed to the difference in their targets on endothelial cells. To reveal the difference, our study focused on their effects on cell signaling pathways. CA is a unique compound because it has various biological effects, especially in the neural system. PA is used for the synthesis of CA [7]. If PA exerts similar effects as CA in the neural system, PA may be used instead of CA. Thus, research is underway, and we have found that PA affects neural cell function similar to CA, but the mechanisms seem to be different (unpublished data). The difference would be related to their mechanisms of antiangiogenic and antilymphangiogenic activities.
Materials and Methods
PA (purity > 99 %), isolated as previously described [1], was dissolved in DMSO to 20 mM as a stock solution, and kept at − 25 °C until use. As positive control inhibitors, CA (purity > 91 %) from Sigma-Aldrich and (−)-epigallocatechin gallate (EGCG; purity > 99 %) from Nacalai Tesque were used for angiogenesis [11] and lymphangiogenesis [19], respectively. The purity of all compounds was determined by HPLC. All animal experiments were approved by the animal experiment committee of Hiroshima University Animal Research Committee; the approval numbers were C11-1 (2011/04/12) and C13–2 (2013/04/11). The ex vivo angiogenesis assay was performed according to a previously reported method [20], [21]. The ex vivo lymphangiogenesis assay was performed according to a previously described method, with some modifications [17]. Rat thoracic ducts were excised from the rats and cut into approximately 1 mm sections under a stereomicroscope. The lymphatic duct rings were embedded in type I collagen gel in a 6-well culture plate as described in the ex vivo angiogenesis assay (Supporting Information). After 30 min to form a gel at 37 °C in a CO2 incubator, 2 mL of RPMI 1640 medium (Gibco) containing 10 % FBS (Lonza), 100 U/mL penicillin, and 100 µg/mL streptomycin (Gibco) were added to the wells. PA or EGCG solution or DMSO was added, and the plates were incubated for 7 days at 37 °C in a CO2 incubator. The images of the lymphatic rings were taken with a digital camera (OLYMPUS DSE330-A system), and the length of the capillaries was measured using Adobe Photoshop software.
Supporting information
A detailed description of the ex vivo angiogenesis assay and in vitro assays using HUVEC and LEC and statistical analysis are available as Supporting Information.
#
#
Acknowledgements
This work was supported in part by grants from JSPS KAKENHI, grant numbers 22580130 and 26450159 (K. M.).
#
#
Conflict of Interest
The authors declare no conflict of interest.
-
References
- 1 Yatagai M, Shirato T, Hayashi Y, Fukuhara N, Takahashi T. Pisiferic acid, a new phenolic diterpene carboxylic acid from Chamaecyparis pisifera S. et Z. J Jpn Wood Res Soc 1978; 24: 267-269
- 2 Kobayashi K, Kuroda K, Shinomiya T, Nishino C, Ohya J, Sato S. Effect of pisiferic acid and its derivatives on cytotoxicity macromolecular synthesis and DNA polymerase alpha of HeLa cells. Int J Biochem 1989; 21: 463-468
- 3 Kang TH, Hwang EI, Yun BS, Shin CS, Kim SU. Chitin synthase 2 inhibitory activity of O-methyl pisiferic acid and 8, 20-dihydroxy-9(11), 13-abietadien-12-one, isolated from Chamaecyparis pisifera . Biol Pharm Bull 2008; 31: 755-759
- 4 Xiao D, Kuroyanagi M, Itani T, Matsuura H, Udayama M, Murakami M, Umehara K, Kawahara N. Studies on constituents from Chamaecyparis pisifera and antibacterial activity of diterpenes. Chem Pharm Bull (Tokyo) 2001; 49: 1479-1481
- 5 Aburai N, Yoshida M, Ohnishi M, Kimura K. Pisiferdiol and pisiferic acid isolated from Chamaecyparis pisifera activate protein phosphatase 2C in vitro and induce caspase-3/7-dependent apoptosis via dephosphorylation of Bad in HL60 cells. Phytomedicine 2010; 17: 782-788
- 6 Aburai N, Yoshida J, Kobayashi M, Mizunuma M, Ohnishi M, Kimura K. Pisiferdiol restores the growth of a mutant yeast suffering from hyper-activated Ca2+-signaling through calcineurin inhibition. FEMS Yeast Res 2013; 13: 16-22
- 7 Tada M, Ohkanda T, Kurabe J. Syntheses of carnosic acid and carnosol, anti-oxidants in Rosemary, from pisiferic acid, the major constituent of Sawara. Chem Pharm Bull (Tokyo) 2010; 58: 27-29
- 8 Aruoma OI, Halliwell B, Aeschbach R, Loligers J. Antioxidant and pro-oxidant properties of active rosemary constituents: carnosol and carnosic acid. Xenobiotica 1992; 22: 257-268
- 9 Kosaka K, Mimura J, Itoh K, Satoh T, Shimojo Y, Kitajima C, Maruyama A, Yamamoto M, Shirasawa T. Role of Nrf2 and p 62/ZIP in the neurite outgrowth by carnosic acid in PC12 h cells. J Biochem 2010; 147: 73-81
- 10 Satoh T, Rezaie T, Seki M, Sunico CR, Tabuchi T, Kitagawa T, Yanagitai M, Senzaki M, Kosegawa C, Taira H, McKercher SR, Hoffman JK, Roth GP, Lipton SA. Dual neuroprotective pathways of a pro-electrophilic compound via HSF-1-activated heat-shock proteins and Nrf2-activated phase 2 antioxidant response enzymes. J Neurochem 2011; 119: 569-578
- 11 Kayashima T, Matsubara K. Antiangiogenic effect of carnosic acid and carnosol, neuroprotective compounds in rosemary leaves. Biosci Biotechnol Biochem 2012; 76: 115-119
- 12 Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med 1995; 1: 27-31
- 13 Alitalo K, Tammela T, Petrova TV. Lymphangiogenesis in development and human disease. Nature 2005; 438: 946-953
- 14 Sugawara T, Matsubara K, Akagi R, Mori M, Hirata T. Antiangiogenic activity of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. J Agric Food Chem 2006; 54: 9805-9810
- 15 Ganesan P, Matsubara K, Ohkubo T, Tanaka Y, Noda K, Sugawara T, Hirata T. Anti-angiogenic effect of siphonaxanthin from green alga, Codium fragile . Phytomedicine 2010; 17: 1140-1144
- 16 Ganesan P, Matsubara K, Sugawara T, Hirata T. Marine algal carotenoids inhibit angiogenesis by down-regulating FGF-2-mediated intracellular signals in vascular endothelial cells. Mol Cell Biochem 2013; 380: 1-9
- 17 Bruyere F, Melen-Lamalle L, Blacher S, Roland G, Thiry M, Moons L, Frankenne F, Carmeliet P, Alitalo K, Libert C, Sleeman JP, Foidart JM, Noel A. Modeling lymphangiogenesis in a three-dimensional culture system. Nat Methods 2008; 5: 431-437
- 18 Cao R, Ji H, Feng N, Zhang Y, Yang X, Andersson P, Sun Y, Tritsaris K, Hansen AJ, Dissing S, Cao Y. Collaborative interplay between FGF-2 and VEGF-C promotes lymphangiogenesis and metastasis. Proc Natl Acad Sci U S A 2012; 109: 15894-15899
- 19 Mineva ND, Paulson KE, Naber SP, Yee AS, Sonenshein GE. Epigallocatechin-3-gallate inhibits stem-like inflammatory breast cancer cells. PLoS One 2013; 8: e73464
- 20 Mori M, Sadahira Y, Kawasaki S, Hayashi T, Notohara K, Awai M. Capillary growth from reversed rat aortic segments cultured in collagen gel. Acta Pathol Jpn 1988; 38: 1503-1512
- 21 Matsubara K, Mori M, Matsuura Y, Kato N. Pyridoxal 5′-phosphate and pyridoxal inhibit angiogenesis in serum-free rat aortic ring assay. Int J Mol Med 2001; 8: 505-508
Correspondence
-
References
- 1 Yatagai M, Shirato T, Hayashi Y, Fukuhara N, Takahashi T. Pisiferic acid, a new phenolic diterpene carboxylic acid from Chamaecyparis pisifera S. et Z. J Jpn Wood Res Soc 1978; 24: 267-269
- 2 Kobayashi K, Kuroda K, Shinomiya T, Nishino C, Ohya J, Sato S. Effect of pisiferic acid and its derivatives on cytotoxicity macromolecular synthesis and DNA polymerase alpha of HeLa cells. Int J Biochem 1989; 21: 463-468
- 3 Kang TH, Hwang EI, Yun BS, Shin CS, Kim SU. Chitin synthase 2 inhibitory activity of O-methyl pisiferic acid and 8, 20-dihydroxy-9(11), 13-abietadien-12-one, isolated from Chamaecyparis pisifera . Biol Pharm Bull 2008; 31: 755-759
- 4 Xiao D, Kuroyanagi M, Itani T, Matsuura H, Udayama M, Murakami M, Umehara K, Kawahara N. Studies on constituents from Chamaecyparis pisifera and antibacterial activity of diterpenes. Chem Pharm Bull (Tokyo) 2001; 49: 1479-1481
- 5 Aburai N, Yoshida M, Ohnishi M, Kimura K. Pisiferdiol and pisiferic acid isolated from Chamaecyparis pisifera activate protein phosphatase 2C in vitro and induce caspase-3/7-dependent apoptosis via dephosphorylation of Bad in HL60 cells. Phytomedicine 2010; 17: 782-788
- 6 Aburai N, Yoshida J, Kobayashi M, Mizunuma M, Ohnishi M, Kimura K. Pisiferdiol restores the growth of a mutant yeast suffering from hyper-activated Ca2+-signaling through calcineurin inhibition. FEMS Yeast Res 2013; 13: 16-22
- 7 Tada M, Ohkanda T, Kurabe J. Syntheses of carnosic acid and carnosol, anti-oxidants in Rosemary, from pisiferic acid, the major constituent of Sawara. Chem Pharm Bull (Tokyo) 2010; 58: 27-29
- 8 Aruoma OI, Halliwell B, Aeschbach R, Loligers J. Antioxidant and pro-oxidant properties of active rosemary constituents: carnosol and carnosic acid. Xenobiotica 1992; 22: 257-268
- 9 Kosaka K, Mimura J, Itoh K, Satoh T, Shimojo Y, Kitajima C, Maruyama A, Yamamoto M, Shirasawa T. Role of Nrf2 and p 62/ZIP in the neurite outgrowth by carnosic acid in PC12 h cells. J Biochem 2010; 147: 73-81
- 10 Satoh T, Rezaie T, Seki M, Sunico CR, Tabuchi T, Kitagawa T, Yanagitai M, Senzaki M, Kosegawa C, Taira H, McKercher SR, Hoffman JK, Roth GP, Lipton SA. Dual neuroprotective pathways of a pro-electrophilic compound via HSF-1-activated heat-shock proteins and Nrf2-activated phase 2 antioxidant response enzymes. J Neurochem 2011; 119: 569-578
- 11 Kayashima T, Matsubara K. Antiangiogenic effect of carnosic acid and carnosol, neuroprotective compounds in rosemary leaves. Biosci Biotechnol Biochem 2012; 76: 115-119
- 12 Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med 1995; 1: 27-31
- 13 Alitalo K, Tammela T, Petrova TV. Lymphangiogenesis in development and human disease. Nature 2005; 438: 946-953
- 14 Sugawara T, Matsubara K, Akagi R, Mori M, Hirata T. Antiangiogenic activity of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. J Agric Food Chem 2006; 54: 9805-9810
- 15 Ganesan P, Matsubara K, Ohkubo T, Tanaka Y, Noda K, Sugawara T, Hirata T. Anti-angiogenic effect of siphonaxanthin from green alga, Codium fragile . Phytomedicine 2010; 17: 1140-1144
- 16 Ganesan P, Matsubara K, Sugawara T, Hirata T. Marine algal carotenoids inhibit angiogenesis by down-regulating FGF-2-mediated intracellular signals in vascular endothelial cells. Mol Cell Biochem 2013; 380: 1-9
- 17 Bruyere F, Melen-Lamalle L, Blacher S, Roland G, Thiry M, Moons L, Frankenne F, Carmeliet P, Alitalo K, Libert C, Sleeman JP, Foidart JM, Noel A. Modeling lymphangiogenesis in a three-dimensional culture system. Nat Methods 2008; 5: 431-437
- 18 Cao R, Ji H, Feng N, Zhang Y, Yang X, Andersson P, Sun Y, Tritsaris K, Hansen AJ, Dissing S, Cao Y. Collaborative interplay between FGF-2 and VEGF-C promotes lymphangiogenesis and metastasis. Proc Natl Acad Sci U S A 2012; 109: 15894-15899
- 19 Mineva ND, Paulson KE, Naber SP, Yee AS, Sonenshein GE. Epigallocatechin-3-gallate inhibits stem-like inflammatory breast cancer cells. PLoS One 2013; 8: e73464
- 20 Mori M, Sadahira Y, Kawasaki S, Hayashi T, Notohara K, Awai M. Capillary growth from reversed rat aortic segments cultured in collagen gel. Acta Pathol Jpn 1988; 38: 1503-1512
- 21 Matsubara K, Mori M, Matsuura Y, Kato N. Pyridoxal 5′-phosphate and pyridoxal inhibit angiogenesis in serum-free rat aortic ring assay. Int J Mol Med 2001; 8: 505-508