Keywords
mouse model - platelets - thromboembolism
Introduction
Animal models allow the investigation of platelets in the contexts of haemostasis,
thrombosis and inflammation while taking into account systemic factors, including
the vascular endothelium, which are not present in isolated platelet preparations.[1]
[2]
[3] The genetic malleability of the mouse makes it the species of choice for many biomedical
researchers. Research involving the use of animals within the European Union (EU)
is regulated by a directive requiring explicit application of replacement (avoidance
of animal use), reduction (use of fewer animals) and refinement (modifying procedures
to reduce severity) of animal procedures (the 3Rs).
Thromboembolic mortality models in which platelet agonists such as collagen, thrombin
and adenosine diphosphate (ADP) are injected into the tail vein of conscious mice
are currently used by significant numbers of research groups to assess platelet function
in vivo.[4]
[5]
[6] These models employ death or hind limb paralysis as endpoints, induce extensive
pain and suffering since anaesthesia is often not used and involve large numbers of
animals ([Table 1]). The scientific value of these models has also been questioned.[7]
Table 1
List of publications using mouse mortality models of thromboembolism in 2016
|
Research groups (2016)
|
Number of animals/group
|
Total number of animals
|
Duration of procedure
|
Platelet agonist
|
Anaesthesia
|
Endpoints
|
|
Assafim[22]
|
Not reported
|
Not reported
|
15 min
|
Thrombin
|
No
|
Death
|
|
Choi[23]
|
10
|
50
|
15 min
|
Thrombin
|
No
|
Paralysis/death
|
|
Huang[24]
|
5
|
15
|
1 h
|
Collagen/adrenaline
|
Yes
|
Death
|
|
Kim[25]
|
15
|
75
|
15 min
|
Collagen/adrenaline
|
No
|
Death
|
|
Park[26]
|
40
|
160
|
15 min
|
Collagen/adrenaline
|
No
|
Paralysis/death
|
|
Misra[27]
|
5 (control)
10 (treatment)
|
25
|
15 min
|
Collagen/adrenaline
|
No
|
Paralysis/death
|
|
Saito[28]
|
3
|
15
|
5 min to 5 days
|
Thrombin
|
No
|
Death
|
|
Lee[29]
|
20
|
Not reported
|
15 min
|
Collagen/adrenaline
|
No
|
Paralysis/death
|
|
Kim[30]
|
10 (control)
16 (treatment)
|
58
|
15 min
|
Collagen/adrenaline
|
No
|
Paralysis/death
|
Note: The table reports a PubMed search list of publications using mortality models
of thromboembolism in 2016 together with the number of animals (total and per group),
platelet agonist, anaesthetic and endpoints used. PubMed keywords: animal/mouse model
thromboembolism, animal/mouse model thrombosis.
Our group has previously developed a refined real-time mouse model of thromboembolism
as an alternative to thromboembolic mortality models.[8] In this model, platelet aggregation is measured directly and in situ in mice under
terminal anaesthesia. The model consists of monitoring 111In-radiolabelled platelets that freely circulate and accumulate in the pulmonary vascular
bed following platelet activation with non-lethal doses of platelet agonists. In vivo
platelet aggregation is measured as increases in platelet-associated counts via external
scintillation probes above the pulmonary region, thus providing the important data
set of quantitative distribution.[8]
[9] The use of general anaesthesia means that painful procedures are avoided and, unlike
mortality models, observations may be directly attributed to platelets.
Real-time radiolabelled platelet monitoring has been shown to be sensitive enough
to identify environmental or nutritional drivers of cardiovascular risk or protection.
For example, our group has shown that platelet aggregation in vivo is enhanced by
diesel exhaust particles or engineered nanoparticles and inhibited by inorganic nitrate/nitrite.[10]
[11]
[12] When applied, the model has led to up to 85% reduction in mouse use compared with
comparable studies using thromboembolic mortality models.[7]
[13]
[14]
Platelet monitoring in vivo requires particular technical skills and handling of radioactive
isotopes along with extensive facilities and equipment. These factors have limited
implementation by groups currently using mortality models of thromboembolism. We therefore
sought novel and refined approaches to study platelet aggregation in vivo that could
be more easily adopted. We now report a non-invasive and more simplistic approach
to study platelet function in vivo based on blood microsampling, with the caveat that
the extent of platelet localization obtained by platelet monitoring in vivo is lacking.
Moreover, since indium oxine is no longer easily available in many parts of Europe,
including the United Kingdom, we characterized indium tropolone for use in real-time
monitoring.
Methods
Mice
Balb/c mice (20–25 g) were purchased from Harlan (Bicester, UK) and had access to
food and water ad libitum. All procedures were conducted under our Home Office Project
License PPL 70/8791 and were approved by the Ethical Review Panel at Imperial College
London, United Kingdom. Protocols were refined in association with the National Centre
for the Replacement, Refinement & Reduction of Animals in Research (NC3Rs)[8] and are reported in accordance with the ARRIVE (Animal Research: Reporting of In
Vivo Experiments) guidelines for reporting experiments involving animals.[15]
Materials
Reagents were purchased from Sigma (Dorset, UK) with the exception of indium oxine
(GE Healthcare, Bucks, UK), indium chloride (Mallinckrodt Pharmaceuticals, Institute
of Nuclear Medicine UCLH, London UK), tropolone (Alfa Aesar, Heysham, United Kingdom)
and collagen (Nycomed Pharma, Unterschleissheim, Germany).
Blood Collection and Platelet Labelling with Indium Oxine or Indium Tropolone
Blood (0.7–1.0 mL) was collected from terminally anaesthetized donor mice (urethane
25% w/v; 10 µL/g) by cardiac puncture into acidified citrate solution (acid citrate-dextrose(ACD);
200 µL). A Ca2+- and Mg2+-free Tyrode's buffer was prepared (CFTS: 138-mM NaCl, 2.6-mM KCl, 5.5-mM glucose,
12-mM NaHCO3 and 0.2-mM NaHPO4) and used to prepare a CFTS/ACD solution (10:1 mixture) with 0.34 µL/mL PGE1 (stock 1 mg/mL). Blood was centrifuged (3 minutes; 300 ×g) to obtain platelet-rich plasma (PRP). CFTS/ACD solution (400 µL) was added to the
remaining red blood cells (RBCs) and centrifuged again (3 minutes; 300 ×g) to acquire additional PRP. The PRP obtained was spun (2 minutes; 200 ×g) to remove contaminating RBCs. Platelets were pelleted (7 minutes; 1,500 ×g) and pooled together by carefully re-suspending them in 1 mL of CFTS/ACD. Pooled
platelets were incubated with 1.8 MBq 111In oxine or 111in tropolone at room temperature for 10 minutes (indium chloride and tropolone 1 µg/µL
were pre-incubated for 10 minutes prior to labelling platelets). Radioactive counts
were measured in samples at this point. Platelets were pelleted (5 minutes; 1,500
×g) and the radioactive supernatant discarded. The surface of the pellet was carefully
washed with CFTS to remove excess radiation and the pellet was then gently re-suspended
in CFTS in a volume of approximately 250 μL per mouse. Radioactive counts were measured
again to calculate percentage labelling efficiency in five independent preparations.
Platelets were left resting for 30 minutes to allow inhibitors to dissipate.
Real-Time Platelet Monitoring In Vivo
Radiolabelled platelets were infused via an exposed femoral vein into anaesthetized
Balb/c mice (urethane 25% w/v; 10 µL/g) and allowed to equilibrate for 20 minutes
as previously described.[8] Platelet aggregation was induced through the intravenous injection of collagen (50
µg/kg) and platelet responses were measured as increases in platelet-associated radioactive
counts in the pulmonary vascular bed. Data were acquired with a Single Point Extended
Area Ratio (SPEAR) probe (eV products, Saxonburg, Pennsylvania, United States) positioned
over the pulmonary vasculature and custom software (Mumed systems, London, United
Kingdom) as previously described.[8]
Blood Sampling via Cardiac Puncture
Anaesthetized (urethane 25% w/v, 10 µL/g) Balb/c mice were injected intravenously
via the femoral vein with increasing doses of collagen (12.5–50 µg/kg) or saline.
Blood was collected by cardiac puncture 1 minute after agonist injection and incubated
with Stromatol (1:100 DF) to measure platelet counts with a haemocytometer (platelets/mL).
Blood Microsampling
Balb/c mice were pre-warmed (37°C) in a surgery recovery box for 10 minutes to facilitate
both microsampling and injections. Animals were anaesthetized with isoflurane (2 minutes)
in the induction box and then transferred to the mask/set-up ([Supplementary Figure S1A] [online only]), maintained under general anaesthesia and ultimately overdosed with
pentobarbital. A small incision was performed with a blade on the distal portion of
the tail vein ([Supplementary Figure S1B] [online only]) and a blood microsample (2 µL) was collected with a pipette (10 μL),
transferred into an Eppendorf tube containing Stromatol (1:100 DF) and mixed to measure
platelet counts (platelets/mL). Bleeding was promptly stopped by putting pressure
on the site of the incision with tissue immediately after blood collection. Collagen
(50 µg/kg), thrombin (65 IU/kg) or ADP (400 μg/kg) were then injected intravenously
in 100 μL volumes via the same tail vein above the incision ([Supplementary Figure S1C] [online only] and https://vimeo.com/192527541). Additional small incisions were performed with a blade 1 and 10 minutes after platelet
activation on the unused tail vein and blood samples (2 µL) collected to measure platelet
counts ([Supplementary Figure S1D] [online only]). In aspirin studies, mice were injected intraperitoneally with vehicle
or aspirin (20 mg/kg) 1 hour prior to platelet activation with collagen (50 µg/kg).
Histology
Balb/c mice were injected intravenously with collagen (50 µg/kg) through the tail
vein as described for the blood microsampling technique. At a time point corresponding
to the maximal response (1 minute), lungs were immediately excised and frozen with
liquid nitrogen. Sections of lung (10 µm) were stained for the platelet-specific surface
antigen CD41 (αIIb) using a specific rat antimouse monoclonal immunoglobulin G (IgG)
antibody (Abcam ab33661, Cambridge, United Kingdom).
Data Analysis and Statistics
The data obtained with the platelet monitoring technique were expressed as percentage
increase in maximal radioactive counts from the baseline recording. All data were
expressed as mean ± SEM (standard error of the mean). Where statistical comparisons
were made, a student's paired t-test or one-way average analysis of variance (ANOVA; repeated measurements) with
Bonferroni post hoc multiple comparison test was used to compare mean values. Data
were considered statistically significant with p-values <0.05.
The labelling efficiency of indium oxine and indium tropolone was calculated as the
percentage of radioactive platelet counts obtained following removal of extracellular
isotope relative to the counts obtained upon initial addition of radioactivity to
platelet suspensions.
Results
Indium Tropolone Efficiently Radiolabelled Isolated Mouse Platelets
Indium oxine and indium tropolone radiolabelled isolated mouse platelets with an efficiency
of 64.4 and 77.8%, respectively ([Table 2]). With both radiolabelling methods, the injection of sub-lethal doses of collagen
(50 µg/kg) caused a rapid increase of platelet-associated radioactivity in the pulmonary
vasculature, which reached a maximal level after approximately 1 minute before returning
to basal levels within 10 minutes ([Fig. 1]).
Table 2
Efficiency of indium oxine and indium tropolone in radiolabelling mouse platelets
|
Indium oxine
|
|
Counts upon initial addition of 111In
|
102,905 ± 4,271
|
|
Counts following platelet labelling with 111In
|
66,158 ± 2,165
|
|
% radioactive labelling
|
64.4 ± 0.0
|
|
Indium tropolone
|
|
Counts upon initial addition of 111In
|
69,333 ± 3,232
|
|
Counts following platelet labelling with 111In
|
55,000 ± 2,733
|
|
% radioactive labelling
|
77.8 ± 3.8
|
Notes: Indium oxine and indium tropolone radiolabelled mouse platelets with an efficiency
of 64.4 and 77.8%, respectively. Efficiency was calculated as percentage of radioactive
counts following platelet labelling procedures compared with radioactive counts upon
initial addition of indium oxine or indium tropolone.
 Fig. 1 Comparison of indium oxine and indium tropolone in a radiolabelled thromboembolic
model of platelet thromboembolism in vivo. Mouse platelets were isolated, radiolabelled
with 1.8 MBq of indium oxine (A, B) or indium tropolone (C, D) and injected intravenously into a donor mouse. A thromboembolic response was induced
via the intravenous injection of collagen and the changes in platelet-associated counts
in the pulmonary vasculature were measured over 10 minutes. Platelet counts rapidly
increased and reached a peak 1 minute after collagen injection and returned to baseline
after 10 minutes. Repeated measures one-way ANOVA (analysis of variance) with Bonferroni
post hoc test; n = 5 (***p < 0.0001).
Fig. 1 Comparison of indium oxine and indium tropolone in a radiolabelled thromboembolic
model of platelet thromboembolism in vivo. Mouse platelets were isolated, radiolabelled
with 1.8 MBq of indium oxine (A, B) or indium tropolone (C, D) and injected intravenously into a donor mouse. A thromboembolic response was induced
via the intravenous injection of collagen and the changes in platelet-associated counts
in the pulmonary vasculature were measured over 10 minutes. Platelet counts rapidly
increased and reached a peak 1 minute after collagen injection and returned to baseline
after 10 minutes. Repeated measures one-way ANOVA (analysis of variance) with Bonferroni
post hoc test; n = 5 (***p < 0.0001).
Platelet Thromboembolism Caused a Measurable Fall in Circulating Platelets
The radiolabelled platelet aggregation response ([Fig. 1]) has previously been shown to be accompanied by a fall in radioactive counts in
areas external to the lung, which then return to basal levels.[8] Prior to devising microsampling techniques, we conducted proof of principle studies
to confirm that platelet accumulation in the lung led to quantifiable reductions in
circulating platelet counts by collecting blood by cardiac puncture in anaesthetized
mice 1 minute after platelet activation with collagen (12.5–50 µg/kg). Collagen at
25 and 50 µg/kg caused a significant drop in platelet counts in blood samples compared
with saline control ([Fig. 2]). We therefore proceeded to refine this technique using microsampling techniques
at multiple time points.
 Fig. 2 Measurement of circulating platelet counts following collagen injection using cardiac
puncture. Anaesthetized mice were injected intravenously (femoral vein) with increasing
doses of collagen (12.5–50 µg/kg) or PBS (phosphate-buffered saline). After 1 minute,
blood was collected by cardiac puncture and incubated with Stromatol. Collagen at
25 and 50 µg/kg caused a significant reduction in platelet counts, 57 and 64%, respectively,
compared with saline control. One-way ANOVA (analysis of variance) with Bonferroni
post hoc test; n = 4 (*p < 0.05).
Fig. 2 Measurement of circulating platelet counts following collagen injection using cardiac
puncture. Anaesthetized mice were injected intravenously (femoral vein) with increasing
doses of collagen (12.5–50 µg/kg) or PBS (phosphate-buffered saline). After 1 minute,
blood was collected by cardiac puncture and incubated with Stromatol. Collagen at
25 and 50 µg/kg caused a significant reduction in platelet counts, 57 and 64%, respectively,
compared with saline control. One-way ANOVA (analysis of variance) with Bonferroni
post hoc test; n = 4 (*p < 0.05).
Changes in Circulating Platelets were Measurable Using Microsampling
Blood microsamples were collected before and after intravenous injection of collagen
(50 µg/kg), thrombin (65 IU/kg) or ADP (400 µg/kg). The data show a significant reduction
in platelet counts 1 minute after injection of collagen ([Fig. 3A]; **p < 0.01) or thrombin ([Fig. 3B]; *p < 0.05) compared with basal platelet counts. After 10 minutes, the counts were restored
for both agonists so that the platelet counts at 10 minutes were not significantly
different to basal counts (p > 0.05; [Fig. 3A], [B]). These results were validated by immunohistochemistry where collagen caused the
accumulation of platelet aggregates, visible as brown staining at ×20 magnification,
in lung tissue; however, this did not occur with the PBS (phosphate-buffered saline)
control ([Fig. 3D]). In contrast, platelet activation with ADP did not show a significant drop in platelet
counts after 1 minute (23.9%; p > 0.05), but after 10 minutes a significant fall in platelet counts was detected
(37.9%; *p < 0.05; [Fig. 3C]).
 Fig. 3 Measurement of circulating platelet counts following agonist administration using
blood microsampling. (A) Anaesthetized mice were intravenously injected with collagen (50 µg/Kg), thrombin
(65 IU/kg), adenosine diphosphate (ADP; 400 µg/kg) or saline. Blood (2 µL) was collected
prior to injection and 1 and 10 minutes after injection by microsampling and platelet
counts determined. After 1 minute, both (A) collagen (n = 5; **p < 0.01) and (B) thrombin (n = 4; *p < 0.05), caused a significant reduction in platelet counts, whereas (C) ADP had no effect at 1 minute although a significant reduction in counts was observed
at 10 minutes. Platelet counts had recovered 10 minutes after injection of both collagen
and thrombin. Repeated measures one-way ANOVA (analysis of variance) with Bonferroni
post hoc test. (D) Sections of mouse lung tissue viewed at ×20 magnification stained for platelet CD41
following injection of saline or collagen showed the formation of aggregates following
collagen administration.
Fig. 3 Measurement of circulating platelet counts following agonist administration using
blood microsampling. (A) Anaesthetized mice were intravenously injected with collagen (50 µg/Kg), thrombin
(65 IU/kg), adenosine diphosphate (ADP; 400 µg/kg) or saline. Blood (2 µL) was collected
prior to injection and 1 and 10 minutes after injection by microsampling and platelet
counts determined. After 1 minute, both (A) collagen (n = 5; **p < 0.01) and (B) thrombin (n = 4; *p < 0.05), caused a significant reduction in platelet counts, whereas (C) ADP had no effect at 1 minute although a significant reduction in counts was observed
at 10 minutes. Platelet counts had recovered 10 minutes after injection of both collagen
and thrombin. Repeated measures one-way ANOVA (analysis of variance) with Bonferroni
post hoc test. (D) Sections of mouse lung tissue viewed at ×20 magnification stained for platelet CD41
following injection of saline or collagen showed the formation of aggregates following
collagen administration.
Aspirin-Inhibited Thromboembolism-Induced Reductions in Circulating Platelets
Aspirin (20 mg/kg) significantly inhibited the reduction in platelet counts from 38.8
to 23.4% compared with vehicle control following platelet activation with collagen
50 μg/kg ([Fig. 4]).
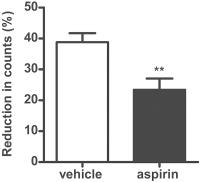 Fig. 4 Measurement of the effect of aspirin on circulating platelet counts following collagen
injection using blood microsampling. Mice were pre-treated for 1 hour with aspirin
(20 mg/kg) or vehicle (n = 6 per group) and then intravenously injected with collagen (50 µg/kg) or saline
under general anaesthesia. Blood (2 µL) was collected before and 1 minute after collagen
injection by microsampling and platelet counts determined. Platelet counts dropped
significantly in the vehicle control group 1 minute after collagen injection, while
treatment with aspirin significantly inhibited this reduction (n = 5). Unpaired one-tailed t-test (**p < 0.01).
Fig. 4 Measurement of the effect of aspirin on circulating platelet counts following collagen
injection using blood microsampling. Mice were pre-treated for 1 hour with aspirin
(20 mg/kg) or vehicle (n = 6 per group) and then intravenously injected with collagen (50 µg/kg) or saline
under general anaesthesia. Blood (2 µL) was collected before and 1 minute after collagen
injection by microsampling and platelet counts determined. Platelet counts dropped
significantly in the vehicle control group 1 minute after collagen injection, while
treatment with aspirin significantly inhibited this reduction (n = 5). Unpaired one-tailed t-test (**p < 0.01).
Discussion
Our group has previously developed a now well-established mouse model of radiolabelled
platelet thromboembolism. The radioisotope required, indium oxine, is increasingly
difficult to source, which led us to develop a new approach for radiolabelling mouse
platelets. Here, we have demonstrated indium tropolone to be an effective alternative
method to radiolabel mouse platelets, which is shown to radiolabel platelets more
efficiently ([Table 2]). With both radiolabelling approaches, we observed changes in radioactive counts
following agonist injection that have been previously shown to correlate with the
formation and consequent resolution of thromboemboli in the dense pulmonary vascular
bed.[8] In the present study, we did not measure platelet counts in other organs or regions.
Previously, however, we have shown platelet accumulation in the pulmonary vasculature
to coincide with a concomitant fall in peripheral regions.[8] This specific accumulation of platelets in the pulmonary vascular bed combined with
histological analysis of lung sections and inhibition of the response by antithrombotic
therapy has led us to conclude previously that our model is driven by platelet aggregation
rather than, for example, adhesion to the vasculature.[8]
Platelet monitoring has the advantage of measuring platelet aggregation in real time
and in situ in the presence of an intact vascular endothelium. This contrasts with
established vascular injury model in which the vascular endothelium is damaged to
induce thrombus formation[16] and which do not functionally isolate the platelet response.[3] Our model has been particularly useful in determining the influence of vascular
mediators such as NO in regulating platelets in vivo, an issue that produced contradictory
findings in vascular injury studies.[17]
[18]
[19]
[20]
[21] Real-time monitoring does, however, require specific skills, resources and facilities.
These issues have proven to be barriers to uptake. We therefore developed blood microsampling
as a refined mouse model to measure platelet aggregation in vivo. Based on published
evidence that platelet activation with collagen causes platelets to accumulate in
the lung with a consequent drop in circulating platelet-associated radioactive counts,[8] blood samples were collected at specific time points following platelet activation
and platelet counts measured. Blood sampling via cardiac puncture was performed under
terminal anaesthesia as proof of principle and confirmed a measurable and significant
reduction in circulating platelet counts in blood samples collected 1 minute after
collagen injection ([Fig. 2]), which correlates with the maximal increase in platelet-associated radioactive
counts detected in the pulmonary vasculature using radiolabelled platelet monitoring
([Fig. 1]). We then refined this approach by developing a blood microsampling method where
multiple small incisions were performed on the tail vein. Similarly to the cardiac
puncture sampling approach, collagen and thrombin, but not ADP, caused a significant
reduction in platelet counts compared with the saline control group which were then
restored over a 10-minute period ([Fig. 3A]). These results were validated by immunohistochemistry where platelet activation
with collagen caused the formation of platelet aggregates in the mouse lung tissue,
whereas this did not occur following saline challenge ([Fig. 3B]). We conclude therefore that the reported reductions in circulating platelets were
due to accumulation of platelet aggregation in the pulmonary vascular bed.
To validate that reductions in platelet counts were driven by platelet activation,
we used the established antiplatelet drug aspirin. Aspirin inhibits the process of
platelet activation by inhibiting the generation of pro-aggregatory thromboxane A2 from cyclooxygenase. The significant effect of aspirin on collagen-mediated changes
in platelet counts links this observation with platelet activation and suggests that
the method may be used to determine the impact of novel antiplatelet compounds or
to investigate other pharmacological or genetic interventions hypothesized to impact
the platelet activation process.
Based on the results presented in this paper, blood microsampling during platelet
thromboembolism represents a refined and non-invasive alternative to thromboembolic
mortality models which avoids pain and suffering in experimental mice and allows the
acquisition of a higher number of samples from each mouse, leading to a considerable
reduction of animal numbers and therefore also of costs. Microsampling does not provide
the continuous data acquisition that arises from real-time monitoring of radiolabelled
platelet accumulation so that the latter remains the gold standard in vivo model.
The importance of real-time monitoring of radiolabelled platelet accumulation is highlighted
by our inability to measure a significant ADP response with microsampling within 1
minute. Microsampling could, however, be implemented by those groups without access
to the expertise or facilities for real-time monitoring and where the full-time course
of the response is not required.
Thromboembolic mortality models employ survival or limb paralysis as endpoints, thus
requiring up to 15 to 20 animals per group to adequately power studies. The 3Rs principles,
now a requirement under EU legislation, require that animals are replaced with non-animal
models wherever possible (replacement), that the number of animals is reduced to the
minimum needed to obtain the results sought (reduction) and that, when used, animal
procedures are refined to minimize their suffering and, thus, that the lowest severity
levels are used (refinement). In compliance with the current EU directive, thromboembolic
mortality models could be replaced with refined real-time monitoring or, where this
is not feasible, microsampling approaches to comply with legislation relating to the
3Rs. Indeed, the application of our platelet monitoring technique has already been
shown to reduce animal numbers by up to 85%[7] and, moreover, the refined microsampling model presented here was able to further
reduce these numbers by around 50%. This additional reduction is achieved by the need
for donor mice in monitoring studies for terminal bleeding to obtain platelets for
radiolabelling and subsequent infusion into recipient mice. As with real-time platelet
monitoring, blood microsampling procedures are conducted at lower severity levels
as animals are kept under terminal anaesthesia throughout procedures and their suffering
would be minimized to constitute a refinement. This contrasts sharply with thromboembolic
mortality studies conducted without anaesthesia or analgesia in which animals are
subjected to considerable pain and suffering.
An additional advantage of both the real-time monitoring and microsampling techniques
reported here is that effects caused by pharmacological or genetic manipulation can
be directly attributed to platelets rather than the less specific endpoint of death
or paralysis that is used in mortality studies. Fewer subsequent experiments are therefore
likely to be required to demonstrate a platelet-mediated effect. In addition, by using
sub-lethal doses of platelet agonists in microsampling studies, we suggest that greater
sensitivity will be achieved compared with mortality studies since effects are tested
at submaximal agonist doses.
In conclusion, we report a simplistic method for assessing platelet thromboembolism
via microsampling. This method provides an alternative to thromboembolic mortality
approaches where the gold standard model, real-time monitoring, is not feasible allowing
scientists to demonstrate compliance with EU legislation concerning the 3Rs.
What is known about this topic?
-
Mouse models of thromboembolic mortality are used worldwide and inflict considerable
suffering and pain. According to European Union (EU) legislation, animal procedures
must be conducted in the context of replacement, refinement and reduction.
-
Platelet monitoring was developed as a refined mouse model of thromboembolism and
measures real-time increases in radioactive-associated platelet counts in the pulmonary
vasculature following the induction of a thromboembolism. This is associated with
a fall in platelet counts in the peripheral vasculature.
-
Real-time platelet monitoring represents a refinement of mortality models since procedures
are conducted under general anaesthesia and considerable reductions in animal numbers
are achieved. However, the technique requires work with radiation, suitable facilities
and particular training.
What does this Paper add?
-
We developed a refined mouse model of thromboembolism based on blood microsampling
where the fall in circulating platelet counts during thromboembolism was measured
and shown to be associated with the accumulation of platelets in the pulmonary vasculature.
This procedure is conducted under general anaesthesia and allows the collection of
three samples from a single mouse.
-
Blood microsampling drastically reduces animal numbers and delivers data by using
a simplistic procedure that does not require work with radiation and that can be easily
implemented by research groups currently using mortality models.
-
An alternative procedure to radiolabel mouse platelets was developed whereby indium
tropolone was used as opposed to indium oxine since the latter is no longer commercially
available in the United Kingdom.




